|
|
Ботаніка Електронний
посібник |
||
|
|
ГЕНЕРАТИВНІ ОРГАНИ. РОЗМНОЖЕННЯ РОСЛИН |
||
|
2.2.
Мікро- і мегаспорогенез. Запилення і запліднення 2.2.1. Розвиток пиляка. Мікроспорогенез.
Утворення чоловічого гаметофіта. 2.2.2. Розвиток насінного зачатка. Мегаспорогенез.
Утворення і розвиток жіночого гаметофіта. 2.2.4. Запліднення. Подвійне запліднення квіткових
рослин. 2.2.5. Апоміксис: партеногенез, апогамія, апоспорія. 2.2.1. Розвиток пиляка.
Мікроспорогенез. Утворення
чоловічого гаметофіта Тичинки
утворюються з меристематичних горбочків квітколожа. Спочатку із цих горбочків
формується пиляк, а потім тичинкова нитка. Через неї і в’язальце проходить
один провідний пучок. У пиляку з меристематичних тканин виникають постійні.
Перші клітини пиляка активно діляться, а пізніше диференціюють. Зовні
утворюється одношаровий епідерміс. Клітини
субепідермального шару, який лежить під епідермісом, тангентально діляться і
з їх внутрішніх шарів формуються клітини археспорія,
а з периферичних клітин утворюється стінка пиляка, яка диференціюється на
три шари: зовнішній – ендотецій,
середній – дегенеруючих клітин і
внутрішній – вистилаючий, або тапетум. Ендотецій формується безпосередньо під епідермісом. Для його клітин характерне
нерівномірне потовщення оболонок. Ендотецій сприяє розкриванню гнізд пиляка
для вивільнення пилку. Під зовнішнім шаром
лежить шар дегенеруючих клітин,
який складається з кількох рядів живих клітин, що утворюють замкнене коло.
Клітини цього шару здавлюються і руйнуються, тому після визрівання пилку його
немає. Тапетум – найбільший внутрішній шар, який безпосередньо прилягає до гнізд з
мікроспорами. Він складається з живих великих, багатоядерних (по 2–4 ядра)
тонкостінних клітин. Це фізіологічно дуже активна тканина, її клітини містять
ферменти, гормони і поживний матеріал, що використовується в процесі
мікроспорогенезу. У одних рослин клітини тапетума руйнуються повністю і йдуть
на живлення мікроспор, а у інших – не руйнуються.
Рис. 2.2.1.1. Будова пиляка в поперечному
розрізі: А –
пиляк з пилковими гніздами; Б – гніздо пиляка; В – пиляк після розкривання В пиляках тичинок
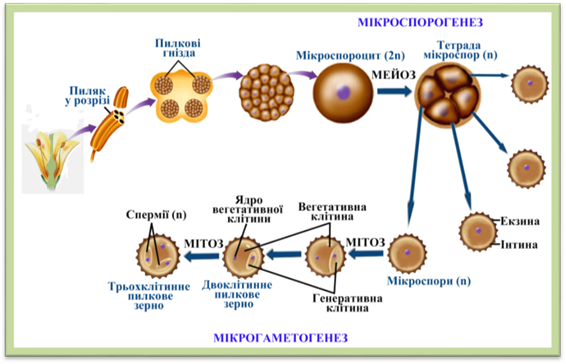
паралельно з диференціацією клітин стінок пиляка відбувається мікроспорогенез – процес утворення
мікроспор. Клітини археоспорія, що розміщуються в центрі гнізда пиляка, мітотично
діляться і перетворюються в диплоїдні
(2п) мікроспороцити, або материнські
клітини мікроспор. Кожен мікроспороцит діляться мейозом і формує тетраду гаплоїдних (п) мікроспор. Тетради
розпадаються на окремі мікроспори до моменту зрілості і відкриття пиляка. У деяких рослин, наприклад у рогоза, тетради не розпадаються на окремі мікроспори. Утворені мікроспори
мають ядро і дві оболонки: зовнішню – екзину
й внутрішню – інтину. У кожному
пиляку звичайно виникають чотири гнізда з мікроспорами. Гаплоїдні
мікроспори, що виникли в гніздах пиляку, звичайно до розкриття пиляків
починають ділитись способом мітозу без цитокінезу (без утворення оболонки між
клітинами). Внаслідок першого поділу, що відбувається під захистом оболонок
мікроспори, виникає проросла мікроспора
з двома неоднаковими голими клітинами. Більшу з них називають клітиною пилкової трубки, нерідко вегетативною, а меншу – генеративною. У деяких рослин
генеративна клітина, знаходячись в пиляку, ще раз поділяється способом мітозу
і дає два спермії. В інших рослин
спермії утворюються пізніше. Пророслу мікроспору (двоклітинну або
триклітинну) вже називають чоловічим
гаметофітом, або пилинкою (пилковим зерном), і служить вона для
запилення. Таким чином, чоловічий гаметофіт квіткових рослин досяг найвищого
ступеня спрощення: він немає проталіальних клітин, антеридія і складається
всього з двох клітин, одна з яких, ділячись, утворює дві безджгутикові
чоловічі гамети – спермії.
Рис. 2.2.1.2. Схема формування чоловічого
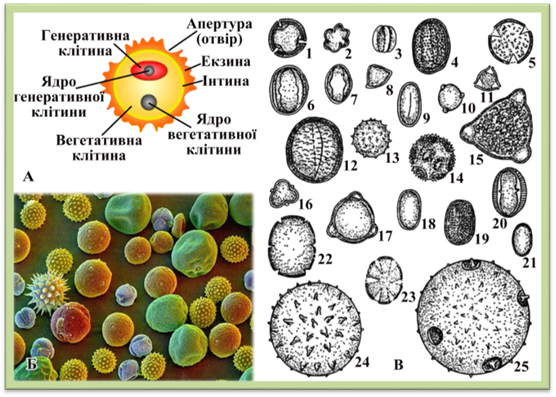
гаметофіту Пилинка містить у
собі певний запас поживних речовин (цукор, крохмаль, олію тощо). Зовні вона має
дві оболонки: зовнішню товсту, часто кутинізовану, – екзину і внутрішню тонку – інтину,
що складається переважно з пектинових речовин. Екзина буває вкрита
потовщеннями у вигляді виростів, горбочків, шишечок, сіточки і має не
потовщені ділянки – апертури
(отвори), крізь які випинається інтина під час проростання пилкової трубки.
Апертури мають вигляд пор або борозен. Форма пилинок у різних видів різна:
куляста, овальна, трикутна, паличкоподібна тощо. Забарвлення пилинок
найчастіше жовтувате чи оранжеве, але буває коричневим, червонуватим,
синюватим тощо. У квітці деяких рослин утворюються тисячі і навіть мільйони
пилкових зерен. Розміри пилинки 0,008 – 2 мм.
Рис. 2.2.1.3. Пилкові зерна: А –
схема будови пилкового зерна; Б – пилкові зерна під електронним мікроскопом;
В – пилок різних рослин: 1 –
липи; 2,3 – фацелії; 4 – гречки; 5 – маку; 6 – конюшини червоної; 7 –
конюшини білої; 8 – акації; 9 – еспарцету; 10 – берези; 11 – ліщина; 12 –
берізки; 13 – соняшника; 14 – кульбаби; 15 – зніту; 16 – верби; 17 – огірка;
18 – медунки; 19 – гірчиці; 20 – волошки; 21 – суріпиці; 22 – будри; 23 –
шавлії; 24 – бавовника; 25 – гарбуза У водному
середовищі пилинка легко набухає і тріскається, в зв’язку з чим квітки мають
багато різних пристосувань, які захищають пилок від дощу. Це
поникаючі квітки (конвалія, брусниця) або суцвіття
(робінія), закривання в похмуру або дощову
погоду квіток (горицвіт, сон розкри́тий) та суцвіть – кошиків (у айстрових), розміщення квіток під захистом листків (липа, розрив-трава) або розміщення
тичинок під пелюстковидними лопатями приймочок (півники) тощо.
video не поддерживается вашим
браузером. Чоловічий гаметофіт 2.2.2. Розвиток
насінного зачатка. Мегаспорогенез. Утворення і розвиток жіночого гаметофіта Насінний зачаток
виникає на плаценті у вигляді напівкулеподібного горбочка, який спочатку
складається з однорідних меристематичних клітин. Через деякий час поблизу
верхівки насінного зачатка з’являється одна або декілька археспоріальних
клітин, які розвиваються з субепідермального шару. Приблизно в цей час біля
основи насінного зачатка виникають інгументи у вигляді одного або двох
кільцевих валиків. Найчастіше в
насінному зачатку під пилковходом одна із субепідермальних клітин нуцелуса – археспоріальна клітина – починає
збільшуватися, а потім поділяється способом мейозу. Виникає тетрада
(четвірка) гаплоїдних (n) мегаспор, які розміщуються здебільшого лінійно. На
цьому мегаспорогенез закінчується. Одна з клітин тетради збільшується,
набуваючи мішкоподібної форми, і стискає три інші клітини, які поступово
розчиняються.
Рис. 2.2.2.1. Схема мегаспорогенезу та
мегагаметогенезу (утворення жіночого гаметофіту) Далі мегаспора
проростає і починається розвиток жіночого гаметофіта. У більшості квіткових
рослин жіночий гаметофіт розвивається з однієї мегаспори шляхом трьох послідовних
мітотичних поділів. Такий тип утворення жіночого гаметофіта називається моноспоричним. Його розвиток
відбувається таким чином. Внаслідок першого поділу ядра мегаспори утворюються
два ядра, які розходяться до полюсів (мікропілярного і халазального) витягнутої
мегаспори, а між ними утворюється велика вакуоля. Далі кожне з цих двох ядер
ділиться синхронно ще двічі і таким чином на кожному полюсі утворюються по
чотири ядра. Це восьмиядерна стадія розвитку жіночого гаметофіта. Таким чином
з мегаспори утворюється восьмиядерний
зародковий мішок (цитокінезу або формування клітинних оболонок під час
поділу тут не буває). Від кожної з двох
полярних четвірок по одному ядру відходить у центральну частину зародкового
мішка. Ці полярні ядра зближуються або зливаються, внаслідок чого утворюється
диплоїдне ядро, яке називається центральним, або вторинним ядром зародкового мішка. Зародковий мішок стає
семиядерним. Центральне ядро покривається цитоплазмою і стає центральною клітиною зародкового мішка
(іноді полярні ядра зливаються пізніше). Біля мікропілярного кінця
зародкового мішка утворюється так званий яйцевий апарат з трьох клітин, які
виникли з трьох ядер, навколо яких сконцентрувалась цитоплазма. Одна з цих
трьох клітин стає яйцеклітиною, або
жіночою гаметою, а дві інші – синергідами (допоміжними клітинами). На халазальному
кінці зародкового мішка виникають три клітини – антиподи. Утворений зародковий мішок із сімома голими клітинами
тепер готовий до процесу запліднення. Цей тип утворення зародкового мішка
трапляється найчастіше, але в природі бувають й інші типи його розвитку.
video
не поддерживается вашим браузером Жіночий гаметофіт 2.2.3. Запилення.
Способи запилення. Пристосування, що запобігають самозапиленню в двостатевих
квітках Запилення
– процес перенесення пилку з пиляка тичинки на приймочку маточки квітки.
Відбувається у рослин під час цвітіння. Розрізняють два типи запилення: самозапилення і перехресне запилення. Самозапилення відбувається
тільки в двостатевих квітках, коли пилок потрапляє на приймочку маточки цієї
самої квітки. Самозапилюються такі
рослини як горох,
квасоля, помідори, бавовник, льон, пшениця, ячмінь, овес
тощо.
Рис. 2.2.3.1. Самозапильні культурні рослини: 1 – овес; 2– просо; 3–рис; 4 – пшениця; 5 – ячмінь; 6 – льон; 7 – горох; 8 – помідори Самозапильні та самозаплідні рослини
називають автогамними. Є автогамні рослини (арахіс, копитняк, розрив-трава,
фіалка, квасениця,
деякі види проса), які мають квітки, що зовсім не
розкриваються, їх називають клейстогамними.
Рис. 2.2.3.2 .Клейстогамні квітки: 1 – фіалки; 2 – квасениці Рослини, які при самозапиленні дають плоди і насіння, називають самофертильними, або самоплідними. Під
час самозапилення добре зберігаються сортові особливості. Але, як зазначав ще
Ч. Дарвін, самозапилення може призвести і до виродження. Самозапилення
обмежує пристосованість рослинних організмів до умов зовнішнього середовища і
тому не сприяє розвитку виду. Перехресне запилення –
більш прогресивний, основний тип запилення квіткових рослин, при якому у
потомстві виявляються спадкові ознаки обох батьків, завдяки чому
відкриваються широкі можливості у пристосуванні до різних умов існування. Суть перехресного запилення в тому, що
пилок з однієї квітки переноситься на приймочку маточки іншої квітки в межах
однієї рослини (гейтоногамія, або сусіднє запилення), або іншої того
самого виду (ксеногамія, або власне перехресне). За способом перенесення пилку перехресне
запилення здійснюється в природі різними шляхами: вітром (анемофілія), комахами (ентомофілія), водою (гідрофілія), птахами (орнітофілія)
тощо.
Рис. 2.2.3.3. Способи перехресного запилення : 1 – птахами; 2 –
комахами; 3 – кажанами; 4 – вітром; 5 – водою Анемофілія. До вітрозапильних
рослин належить близько 10% всіх покритонасінних рослин. Квітки анемофільних рослин
характеризуються відсутністю, або редукцією оцвітини, утворенням великої
кількості дрібних квіток, що не мають запаху, забарвлення і нектару. Пилок
численний, легкий, сипучий. Серед вітрозапильних рослин частіше трапляються
роздільностатеві квітки, зібрані у волоті, колоски, сережки. Більшість
вітрозапильних рослин зацвітає рано навесні, частіше до розвитку до розвитку
листків, або одночасно з ними. До вітрозапильних рослин належать злакові, осокові, березові, букові, горіхові.
Рис. 2.2.3.4. Вітрозапильні
рослини: 1 – осока; 2 – береза; 3 – бук; 4 – подорожник; 5 – горіх; 6 – кропива; 7 – жито; 8
– кукурудза; 9 – хміль Ентомофілія. Комахи відіграли велику
роль не тільки в еволюції квітки, її походженні. Перші покритонасінні рослини
мали двостатеві ентомофільні квітки, які приваблювали комах пилком. Пізніше в
квітках з’явилися нектарники і нектар став додатковим фактором приваблювання
комах. Добре відомо, що комахи відвідують квітки для збору їжі, але інколи
вони, знаходять там притулок, відкладають яйця тощо. Пилок – найважливіше
джерело їжі, яке комахи знаходять у квітці. Він містить білки, жири,
вуглеводи, вітаміни, ферменти, мінеральні елементи. У комахозапилювальних
рослин пилок, як правило, крупний, звичайно з нерівною поверхнею, часто
клейкий. Ці властивості полегшують захват і перенесення його комахами. Крім
того комахи живляться нектаром, що є водним розчином цукрів.
Рис. 2.2.3.5. Бджола обсипана пилком Крім
їжі, запилювачів приваблює також забарвлення віночка і запах. Комахозапильні
рослини мають яскраво забарвлений віночок, а якщо квітки дрібні то вони
зібрані в добре помітні суцвіття (китиці, щитки, зонтики).
Рис. 2.2.3.6. Комахозапильні
рослини: 1 – мак; 2 – соняшник; 3 –
калина; 4 – яблуня; 5 – гречка; 6 – конюшина; 7 – огірок; 8 – липа;
9 – ромашка Багато комах добре орієнтуються на запахи. Так, квітки,
що запилюються бджолами, мають приємний аромат, зумовлений сполуками
бензольного типу. Аміноїдні запахи, зумовлені наявністю амінів приваблюють і жуків (горобина, глід, барбарис). Ароматичні сполуки, виділяються
пелюстками, тичинками, нектарниками або спеціальними багатоклітинними
залозками і поширюються потоками повітря. Рослини, квітки яких, виділяють нектар, називаються медоносними. До них належать представники родини розоцвітих, бобових, гарбузових, губоцвітих, зонтичних та ін. Гідрофілія спостерігається у
водних рослин. Типовими гідрофільними
рослинами є кушир, різуха, валіснерія, морська трава. Характерною
їх ознакою є утворення негроскопічного пилку, який не гине у воді. Пилок течією переноситься на приймочки.
Рис. 2.2.3.7. Гідрофільна
рослина кушир занурений з чоловічою та жіночою квіткою Орнітофілія, тобто запилення птахами характерне для
тропіків зрідка для субтропічних областей. Орнітофільними
рослинами є деякі види евкаліптів, кактусів, алое. Вони приваблюють колібрі,
нектарниць. Для квіток, які запилюють птахи, характерне яскраве забарвлення і
велика кількість пилку.
Рис. 2.2.3.8. Запилення
птахами (колібрі) Зрідка рослини запилюються кажанами, що властиво
тропікам Азії і Америки. Це переважно дерева, ліани, рідше кущі, трави з
великими квітками і міцними пелюстками. Перехресне запилення у ході еволюції виявилось найбільш
прогресивним. При перехресному запиленні у потомстві виявляються спадкові
властивості обох батьків, завдяки чому відкриваються широкі можливості у
пристосуванні до умов існування. Штучне запилення – запилення, яке
здійснює людина. Цей вид запилення
широко застосовують у плодовому і декоративному садівництві, овочівництві, лісовому господарстві і селекційній роботі. При цьому пилок
штучно переносять із тичинок на приймочку маточки запилюваної рослини.
Штучним може бути як самозапилення, так і перехресне запилення. Штучне запилення широко застосовується людиною для
підвищення урожаю і отримання нових сортів рослин. При цьому для перенесення
пилку на приймочку маточки використовують різні способи. Так, у кукурудзи, яка має одностатеві квітки, пилок
збирають, струшуючи верхівки волоті чоловічих квіток у паперові мішечки. Потім зібраний пилок
пензликом переносять на жіночі квітки.
При штучному запиленню соняшнику надягають
рукавичку на руку, знімають нею пилок з однієї рослини і переносять на другу. Можна це зробити притисканням двох
кошиків один до одного. Для отримання нових сортів рослин з двостатевими
квітками необхідна підготовка до штучного запилення. Перш за все із квіток
рослини, обраної за материнську, ще в пуп’янку вилучають пиляки і захищають
ці квітки марлевими або паперовими мішечками від потрапляння пилку. Через 2–3
дні, коли пуп’янок розкриється, наносять на приймочку маточки пилок іншого
сорту чистим пензликом. Пристосування, що
запобігають самозапиленню. Щоб уникнути самозапилення в
перехреснозапильних рослин виробились спеціальні пристосування. Найпоширеніші
дихогамія, гетеростилія, самобезплідність. При дихогамії у квітці відбувається
неодночасне дозрівання пиляків і підготовка приймочок маточки до прийому
пилку. У одних рослин (у губоцвітих, бобових, зонтичних, айстрових, дзвоникових, гвоздикових) пиляки дозрівають
раніше, а приймочки не готові до сприймання пилку. Це явище називають протерандрією. У інших
рослин (хрестоцвітих, розоцвітих, пасльонових, барбарисових) раніше дозріває
приймочка, а пиляки ще не дозріли і не розкрились. Це явище називають протерогінією. При гетеростилії (різностовпчатості)в межах одного
виду одні квітки мають тичинки з довгими тичинковими нитками і маточки з короткими
стовпчиками, інші, навпаки, мають тичинки з короткими тичинковими нитками і
маточки з довгими стовпчиками. Відповідно розміщені і пиляки – в одних
квітках нижче, а в інших – вище власної приймочки, тобто вони завжди бувають
на рівні положенню приймочки в квітках іншої форми. Явище
гетеростилії спостерігається
у квітках первоцвітів, незабудок, гречки, медунок та ін.
Рис. 2.2.3.9. Гетеростилія у
первоцвіта звичайного: 1 – довгостовбчата
форма; 2 – короткостовбчата форма Вчені встановили, що запилення має добрі результати,
коли пилок з квіток з довгими тичинками потрапляє на приймочки маточок, що
мають довгі стовпчики, і, навпаки, пилок з квіток з короткими тичинками запилить
коротко стовпчасті маточки. Таке запилення називають законним (легітимним).
Самозапилення в названих квітках буде незаконним
(ілегітимним), воно позитивних
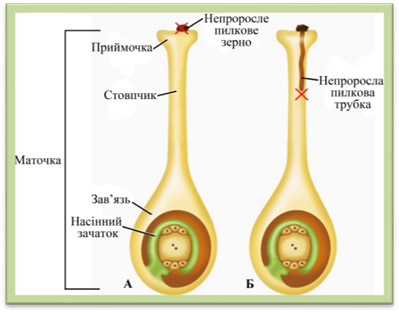
наслідків не дає. Самобезплідність, або самостерильність полягає в тому, що свій пилок, потрапивши
на приймочку маточки, не викличе запліднення. Причини можуть бути різні. В
одних випадках пилкова
трубка не досягає насінного
зачатка, в інших – маточки виділяють
речовини, які перешкоджають проростанню пилкової трубки (у жита, моркви, буряку).
Рис. 2.2.3.10. Схеми
самобезплідності: А – пилкове зерно не проростає на приймочці маточки; Б – пилкова
трубка не досягає насінного
зачатка 2.2.4. Запліднення.
Подвійне запліднення квіткових рослин Потрапивши на
приймочку маточки, пилкове зерно починає поростати. Цьому сприяють речовини,
які виділяють приймочки, відповідні температура, вологість та інші умови
зовнішнього середовища. В процесі проростання вміст пилкового зерна
випинається крізь пору екзини у вигляді тонкої трубочки, вкритої лише інтиною.
Ця пилкова трубка починає видовжуватись і проникає всередину стовпчика. Тут
вона просувається по спеціальному каналу або по міжклітинниках; потім досягає
зав’язі і прямує до мікропіле насінного зачатка. Ріст пилкової трубки
пов'язаний з обміном речовин, що відбувається між нею та внутрішніми
клітинами. У більшості рослин
пилкова трубка проникає
до насінного зачатка крізь
мікропіле (порогамія), у берези, вільхи і деяких інших
– крізь халазу (халазогамія), у
в’яза – крізь інгументи
(мезогамія).
Потім пилкова трубка проникає до зародкового мішка. За період росту всередині
маточки вміст пилкової трубки зазнає певних змін. Вегетативне ядро поступово
розсмоктується, його речовини використовуються на ріст трубки. Генеративне
ядро поділяється способом мітозу на дві чоловічі статеві клітини – спермії (у деяких рослин це
відбувається раніше, ще у непророслій пилинці). При стиканні з зародковим
мішком відбувається розчинення стінки пилкової трубки, а увійшовши до
зародкового мішка, вона розривається, і її вміст виливається на одну з
синергід, яка гине. З двох сперміїв, які звільнились, один прямує до жіночої
клітини – яйцеклітини і зливається
з нею. З диплоїдної зиготи, що виникла, утворюється зародок. Другий спермій зливається з центральною диплоїдною
клітиною зародкового мішка, і триплоїдна клітина, яка утворилась, пізніше
сформує живильну тканину – ендосперм.
Рис. 2.2.4.1. Схема подвійного
запліднення і проростання пилкової трубки у квіткових рослин У цьому злитті двох сперміїв з двома клітинами зародкового мішка полягає суть подвійного запліднення, яке відкрив у 1898
році С.Г. Навашин.
Рис. 2.2.4.2. С. Г. Навашин Після подвійного запліднення
синергіди й антиподи відмирають. Насінний зачаток розростається і
перетворюється в насінину.
Інгументи, видозмінившись, дадуть шкірку
насінини. Зав’язь (а іноді в цілому квітка) перетвориться в плід. Із стінок зав’язі сформується оплодень. Якщо в клітинах нуцелуса
накопичуються поживні речовини, то вони утворять особливу тканину – перисперм. З моменту потрапляння
пилинки на приймочку маточки до процесу запліднення у різних рослин проходить
різний час – від 20–30 хв. до кількох діб. Подвійне
запліднення властиве тільки квітковим рослинам. 2.2.5. Апоміксис:
партеногенез, апогамія, апоспорія Апоміксисом називають різноманітні випадки розвитку зародка без запліднення. У цьому
випадку при утворенні зародкового мішка мейоз не відбувається і всі його
клітини диплоїдні. Диплоїдна
яйцеклітина нередукованого жіночого гаметофіта може дати початок зародку. Це
явище називається партеногенезом.
Розвиток зародка з інших клітин жіночого гаметофіта (синергід, антипод)
називається апогамією, а розвиток
зародка з частин спорофіта (інгументів, нуцелуса) – апоспорією. З апоспорією пов’язана поліембріонія – багатозародковість насінини. Запитання і завдання для самоконтролю 1. Що таке мікроспорогенез? Де і як він відбувається? 2. Що ви знаєте про розвиток і будову чоловічого
гаметофіта? 3. У чому полягає принципова відмінність між пилком і
мікроспорою? 4. Де і як відбувається мегаспорогенез? 5. Як відбувається розвиток жіночого гаметофіта? 6. Яка його будова зародкового мішка? 7. Що таке запилення? Які є способи запилення в природі? 8. Які є способи самозапилення? В чому полягає їх суть? 9. Як може здійснюватись перехресне запилення? 10. Які пристосування є у рослин до запилення вітром,
комахами, водою, птахами? 11. Які є пристосування у
перехреснозапильних рослин, що запобігають самозапиленню в двостатевих
квітках? 12. Що таке штучне
запилення? Для чого його застосовують? 13. Як відбувається подвійне запліднення? Які зміни
відбуваються в насінному зачатку в результаті подвійного запліднення? 14. Які бувають способи утворення зародка без запліднення? |
|||