|
|
Ботаніка Електронний
посібник |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
ЦИТОЛОГІЯ |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
1.2.4. Хімічні видозміни клітинної оболонки 1.2.6. Утворення міжклітинників Походження
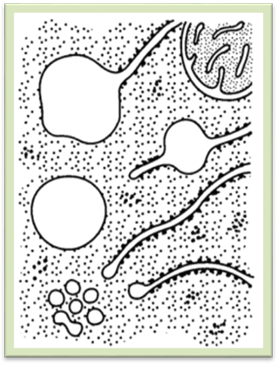
вакуолей. Вакуолі (лат. vacuus – порожнина)
– це порожнини в цитоплазмі, оточені мембраною, що називається тонопластом та заповнені рідиною – клітинним соком (рис. 1.2.1.1). У рослинних
клітинах вакуолі переважно займають понад 30 % об’єму, а в окремих випадках
навіть до 90 %. Рис 1.2.1.1. Рослинна клітина з центральною вакуолею Число і розмір вакуолей у клітині
змінюється з її віком. У молодих клітин виникає багато дрібних вакуолей –
провакуолей. Вони з’являються у вигляді мікропухирців, що відокремлюються від
ендоплазматичної сітки (рис. 1.2.1.2).
Рис 1.2.1.2.
Виникнення вакуолей із елементів ендоплазматичної сітки шляхом утворення локальних здуттів з наступною їх ізоляцією (зверху і в центрі) і відчленуванням пухирців з наступним їх злиттям (знизу) З нагромадженням клітинного соку
провакуолі збільшуються в розмірах і зливаються між собою. У клітин, що
старіють, вакуолі зливаються в одну центральну вакуолю, яка відтісняє
протопласт до периферії клітини (рис. 1.2.1.3).
Рис 1.2.1.3. Розвиток вакуолей в рослинній клітині (1–5) Вода до вакуолі надходить крізь
тонопласт завдяки осмосу, що спричинений вищою концентрацією розчину
клітинного соку. Речовини до вакуолі надходять шляхом активного транспорту,
або ж в мембранних пухирцях, які найчастіше є похідними комплексу Гольджі. Функції
вакуолей різноманітні. По-перше, вони, утримуючи клітину в стані
тургору, виконують разом з клітинною оболонкою опорну функцію. По-друге,
містять гідролітичні ферменти, що розщеплюють складні сполуки. По-третє,
вакуолі накопичують у собі
певні речовини. Серед них є запасні
поживні речовини. Так у насінні квасолі і гороху вакуолі містять
велику кількість білків. Коли насіння проростає білки розщеплюються
ферментами, а амінокислоти транспортуються через тонопласт у циптоплазму.
Також вакуолі можуть зберігати пігменти, такі як антоціанін, зокрема у
пелюстках для приваблення запилювачів. Ці органели також можуть містити
речовини з неприємним смаком або отруйні, такі, як алкалоїди або глікозиди, що
відлякують тварин і захищають рослини від поїдання. Клітинний
сік, його хімічний склад. Основним компонентом клітинного соку
є вода, в якій у вигляді розчину або колоїдів знаходяться різні речовини:
солі, цукри, органічні кислоти та інші розчинні з'єднання, білки і навіть
жирові сполуки. Хімічний склад клітинного соку може змінюватися. Речовини, що
знаходяться в клітинному соку, є або запасними поживними речовинами, які
можуть використовуватися у процесі життєдіяльності, або побічними продуктами
вторинної природи. До запасних речовин належать вуглеводи. Найчастіше
вуглеводи клітинного соку представлені моносахаридами – глюкозою, або
виноградним цукром і фруктозою, або
плодовим цукром. Фруктоза солодша ніж глюкоза. У великій кількості ці цукри
знаходяться в плодах винограду, яблук, груш, слив, персиків, кавунів, у
стеблах сорго і кукурудзи (рис. 1.2.1.4).
Рис 1.2.1.4. Плоди, багаті на моносахариди З дисахаридів найпоширенішою є сахароза, або
тростинний цукор. Багато його у стеблах цукрової
тростини, у коренях цукрових
буряків (рис. 1.2.1.5).
Рис 1.2.1.5. Рослини, багаті на
сахарозу: 1 – тростина цукрова; 2 – буряки цукрові До полісахаридів, які є в клітинному
соку, належить інулін (в клітинах соняшника, топінамбура, цикорію). Інулін відкладається в тканинах
паренхіми підземних органів рослин (рис. 1.2.1.6).
Рис 1.2.1.6. Органи рослин, які містять інулін: 1 – бульби топінамбура; 2 – корінь цикорію До речовин вторинної природи, що
знаходяться у вакуолях, належать алкалоїди, глікозиди, дубильні
речовини і пігменти клітинного соку. Алкалоїди – складні
нітрогеновмісні органічні сполуки лужної реакції переважно рослинного
походження. Вони гіркі на смак, фізіологічно і фармакологічно дуже активні,
часом отруйні або діють як наркотики, не
використовуються в процесах метаболізму. При реакції з кислотами утворюють
солі. Більшість алкалоїдів у чистому вигляді – кристали, а
Рис 1.2.1.7. Рослини, які
містять алкалоїди: 1 – мак снотворний; 2 – беладонна звичайна; 3 – тютюн
звичайний; 4 – ефедра двоколоса; 5 – чистотіл звичайний; 6 – чай китайський Глікозиди – природні органічні речовини, сполуки складної будови, дуже поширені
в рослинному світі; з погляду У рослинах
(рис. 1.2.1.8) найчастіше
зустрічаються такі глікозиди: амигдалин – у кісточках
гіркого мигдалю, абрикоса, персика, сливи, вишні, який розпадається з утворенням дуже отруйної синильної кислоти; синігрин – у гірчиці, хрону; дигоксин – у наперстянки; лінамарин – у льону; гесперідин – у цитрусових; кумарин – у пахучому колоску;
Рис 1.2.1.8. Органи рослин, що містять глікозиди: 1 – насінина
мигдалю; 2 – корінь хріну; 3 – плоди ванілі Дубильні
речовини є в клітинах багатьох рослин, зокрема в корі, плодах,
листках, менше в коренях і стеблах. Вони спричиняють терпкий смак плодів груші чи айви, кори дуба чи крушини, чаю чи кавових зерен. За
своєю природою – це збірна група сполук, яка має властивість дубити,
утворюючи осад з колагеном. Дубильні речовини мають вигляд
колоїдного розчину і певною мірою пов'язані з метаболізмом крохмалю, синтезом
і переміщенням цукрів і асимілятів під час фотосинтезу. Серед дубильних речовин поширені таніни – похідні
галової кислоти. Їх багато в корі дуба, сумах, модрині, галах тощо (рис.
1.2.1.9).
Рис 1.2.1.9. Містять таніни: 1 – кора дуба; 2
– гали; 3 – хурма Негідролізовані конденсовані дубильні
речовини клітинного соку називають флобафенами. Використовують дубильні речовини в
шкіряній, хімічній промисловості, а також у медицині. Органічні
кислоти є майже в усіх клітинах і рослинах. Вони знаходяться у
вільному або зв'язаному стані. Наявність органічних кислот і їх солей
зумовлює кислу реакцію клітинного соку. У рослин зустрічаються такі органічні
кислоти: щавелева, яблучна, винна, янтарна, лимонна тощо. Щавелева
кислота дуже поширена і зустрічається у
вигляді кислих солей калію, натрію, кальцію в щавлю, ревені, квасениці, покісниці
тощо (рис. 1.2.1.10).
Рис 1.2.1.10. Рослини, які містять солі щавлевої кислоти: 1 – щавель; 2 –
шпинат; 3 – ревінь; 4 – квасениця Багато яблучної
кислоти, зокрема в плодах яблуні, малини, барбарису, горобини, айви, аличі (рис.
1.2.1.11).
Рис 1.2.1.11. Рослини, плоди яких містять яблучну кислоту: 1 – яблуня; 2 –
алича; 3 – барбарис; 4 – малина У плодах винограду, шовковиці, вишні, помідорів та інших
рослин (рис. 1.2.1.2) є винна кислота. Вона
знаходиться у вільному стані або у вигляді кальцієвих солей.
Рис 1.2.1.12. Рослини, плоди яких містять винну кислоту: 1 – вишня; 2 –
виноград; 3 – помідор; 4 – шовковиця Лимонна
кислота характерна для плодів лимона. Менше її в
плодах смородини, аґрусу, абрикосів, персиків. Янтарна
кислота є в плодах смородини і суниць, саліцилова – малини, бензойна – журавлини, чорниці, брусниці. Мурашина
кислота є
в кропиві, у хвої сосни, аконітова – в аконітів, гліоксилова і піровиноградна
кислоти –
у листках і коренях кукурудзи, гороху, ячменю, гречки; молочна – у листках квасолі, люцерни, квітках жоржини. Наявність органічних кислот у
плодах насамперед захищає рослини від поїдання тваринами, від руйнування їх
грибами і бактеріями. Вони важливі як у свіжих плодах, так і продуктах
переробки. Саме завдяки їм багато плодів набуває дієтичності. Мінеральні
солі в клітинному соку містяться у вигляді солей нітратної
(нітрати), фосфатної (фосфати), хлоридної
(хлориди) та інших
кислот. На нітрати багаті деякі бур'яни (рис. 1.2.1.13), наприклад кропива жалка, щириця
загнута, грицики
звичайні, з культурних рослин – соняшник, картопля, квасоля та ін.
Рис 1.2.1.13. Рослини, які містять нітрати: 1 – кропива; 2 –
грицики; 3 – щириця; 4 – соняшник Фосфати
зустрічаються в молодих наростаючих частинах рослин у вигляді кальцієвих і
калійних солей, фосфатної кислоти. Чимало міститься солей фосфору у гречки, бобових. Хлориди у вигляді
калієвих і натрієвих солей містяться в стеблах і листках петросимонії, солонцю. Пігменти
клітинного соку. Клітинний сік містить пігменти – речовини,
які забарвлюють тканини рослини в яскраво-червоний, фіалковий, синій або
жовтий кольори з різними відтінками. Яскраве забарвлення репродуктивних
органів рослини має важливе значення у процесі запилення квітки, поширення
плодів. Найпоширенішими пігментами є антоціани,
антохлори, антофеїн. Антоціани становлять
фенольну групу і легко руйнуються, розпадаються на глюкозу та
антоцианід. Вміст антоціану спостерігається уже в молодих проростках.
Антоціани змінюють забарвлення залежно від реакції (рН) клітинного соку (рис.
1.2.1.14). Вакуолі, клітинний сік яких характеризується помірною лужною
реакцією, забарвлюються в синій або блакитний кольори, якщо реакція
нейтральна – у фіалковий. У кислому середовищі антоціани червоні різного
відтінку, залежно від ступеня кислотності.
Рис 1.2.1.14. Зміна
забарвлення розчину антоціанів, виділених із червонокачанної капусти, при зміні рН розчину від 1 до 10 (зліва направо) Всі кольори квітів, за невеликим
винятком, зумовлено наявністю антоціанів у вакуолях клітин епідермісу
пелюсток (рис. 1.2.1.15).
Рис 1.2.1.15. Забарвлення пелюсток квітів зумовлене наявністю антоціанів: 1 – у маку; 2 –
у петунії; 3 – у волошок Клітинний сік може змінювати
забарвлення і від фізіологічного стану рослини. Так, віночок рослин з родини шорстколистих буває
рожевим або червоним до запилення квіток і синім після запилення. Тому в
суцвіттях медунки, незабудки можна
одночасно зустріти квітки, що мають різне забарвлення (рис. 1.2.1.16).
Рис 1.2.1.16. Різне забарвлення квіток у суцвіттях: 1 – медунки; 2 –
незабудки Антоціани зумовлюють фіалковий і
червоний кольори плодів вишні, сливи, суниці,
коренеплодів буряка, редиски та ін. (рис.
1.2.1.17). Наявність антоціану в клітинах підвищує активність поглинання
світла. При цьому використовується частина сонячного спектра, яка не
засвоюється зеленими пігментами листка, температура тканин листка
підвищується, що захищає рослину від негативного впливу низьких температур.
Рис 1.2.1.17. Забарвлення зумовлене наявністю антоціанів: 1 – у
коренеплодів буряків; 2– у плодів чорниці; 3 – у плодів суниці Антохлори – жовті
пігменти клітинного соку. За хімічною природою подібні до антоціанів. Золотисто-жовте
забарвлення квіток лядвенцю, льонку, жоржин; плодів лимона, мандарина, грейпфрута зумовлене
наявністю антохлору
Рис 1.2.1.18. Забарвлення пелюсток квітів зумовлене наявністю антохлору: 1 – у жоржин; 2 –
у льонку; 3 – у лядвенцю Антофеїн – пігмент
темно-бурого кольору, зустрічається рідше, ніж попередні. Він забезпечує
темно-буре забарвлення пелюсток і чашолистків у деяких представників орхідних, пелюсток кінських
бобів, садової фіалки тощо (рис.
1.2.1.19).
Рис 1.2.1.19. Темно-буре забарвлення пелюсток квітів зумовлене наявністю
антофеїну: 1 – у кінських
бобів; 2 – у садової фіалки Пігменти часто поєднуються між собою.
Так, у смородини, в
результаті поєднання антоціану з пігментом Включення – це
продукти обміну речовин протопласта, які з'являються і зникають у різні
періоди життя рослини. Включення можуть міститись у вакуолях, клітинній
оболонці, можуть бути зв'язані з протоплазматичними компонентами клітини. До включення належать запасні
речовини та кінцеві продукти обміну речовин, які представлені кристалами, що
утворені деякими речовинами. Запасні
речовини. У результаті фотосинтезу утворюються органічні
речовини, які використовуються як будівельний або енергетичний матеріал для
формування і розвитку органел клітини. Частина з них відкладається про запас.
Найпоширеніші запасні поживні речовини – вуглеводи, білки, жири. Вуглеводи. Найбільш
поширеною запасною речовиною рослин серед вуглеводів є полісахарид – крохмаль
Рис.1.2.2.1 Крохмаль: 1– картопляний; 2 – кукурудзяний; 3 – пшеничний Довгі лінійні молекули крохмалю побудовані із залишків глюкози (рис. 1.2.2.2).
Рис.1.2.2.2. Фрагмент структурної формули крохмалю Розрізняють крохмаль асиміляційний,
або первинний; транзиторний і запасний, або вторинний. Асиміляційний
крохмаль, або первинний, утворюється в процесі фотосинтезу в
хлоропластах з глюкози. У нічний
час під впливом ферментів він
гідролізується до розчинних сахаридів, які потім транспортуються до місць
споживання чи відкладання в запас. Транзиторний
крохмаль утворюється тимчасово на шляхах переміщення сахаридів
до місць відкладання в запас. Такий крохмаль у вигляді тимчасового утворення
відкладається у високорослих деревних порід, ліан та інших рослин. Запасний
крохмаль, або вторинний, формується у лейкопластах у вигляді
крохмальних зерен. Такі лейкопласти називають амінопластами або крохмалеутворювачами. Ділянка амілопласта,
навколо якої відкладається крохмаль, називається ядром, або утворювальним центром. Крохмаль відкладається шарами
навколо утворювального центра у вигляді світлих і темних шарів. Під час
рівномірного накладання шарів навколо утворювального центра формуються
концентричні крохмальні зерна
(наприклад, у зернівці пшениці), під час
нерівномірних нашарувань утворюються ексцентричні крохмальні зерна (наприклад, у бульбі картоплі). Шаруватість крохмальних зерен
пов’язують із різною щільністю та вмістом вологи в них: світлі шари щільніші
й малозволожені, тоді як темні пухкі з більшим вмістом гігроскопічної вологи.
Інтенсивність і потужність їх формування різна протягом дня і ночі. Розрізняють три типи крохмальних
зерен: прості, складні і напівскладні (рис. 1.2.2.3). Прості зерна мають один утворюючий
центр, навколо якого відкладаються шари крохмалю. У складних зерен в одному лейкопласті кілька утворюючих центрів
(два і більше), кожний з яких має власну шаруватістю. У напівскладних зерен також кілька утворюючих центрів з власною
шаруватістю, оточених спільною шаруватість. Всі ці типи крохмальних зерен
можна знайти у клітинах бульб картоплі.
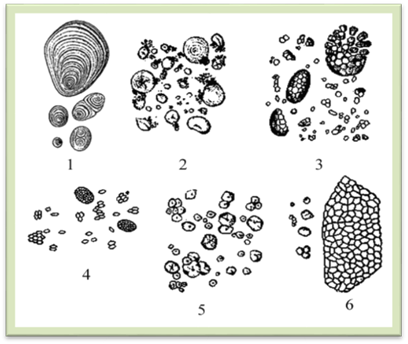
Рис.1.2.2.3. Крохмальні зерна у бульбах картоплі: 1 – просте
ексцентричне зерно; 2– просте концентричне зерно; 3– складне зерно; 4 – напівскладне зерно Кожний вид рослин має властивий йому
тип крохмальних зерен, зумовлений формою, розмірами, структурою та іншими
ознаками (рис. 1.2.2.4). Найменші крохмальні зерна у рису (3–10 мкм),
більші розміри мають крохмальні зерна гороху (50-60 мкм),
а ще більші вони у картоплі (до 145
мкм), і особливо – у петрового хреста (275 мкм).
Рис.1.2.2.4. Крохмальні зерна різних рослин: 1 – картоплі; 2 – пшениці; 3 – вівса; 4 – рису; 5 –
кукурудзи; 6 – гречки Крохмаль
відкладається у різних органах рослин, але найбільше його в насінні,
запасаючих органах, таких як бульби, кореневища, цибулини, в паренхімі
вторинних провідних тканин. Таблиця 1.2.2.1 Вміст крохмалю у
рослинах
Серед полісахаридів відомий інулін, який
знаходиться в колоїдному стані, але може випадати в осад у вигляді
сферокристалів. У бактерій, грибів і синьо-зелених водоростей зустрічається
полісахарид глікоген. У насінних
шкірках багатьох плодів і насінин відкладається в запас геміцелюлоза. Запасні білки. У процесі обміну речовин у клітині виробляються різні
білкові сполуки, зокрема структурні й запасні білки. Серед запасних білків
виділяють аморфні, кристалічні та зернисті.
Аморфний білок утворює глобули, або
безформні маси. Добре відомим аморфним запасним білком є клейковина, яка в ендоспермі зернівки пшениці зв'язана з
крохмалем. Кристалічний білок, як і
крохмаль, об'єднує в собі властивості і кристалів, і колоїдів. Тому його
одиниці ще називають кристалоїдами.
Білкові кристали властиві пластидам, ядру, цитоплазмі, клітинному соку. В
ендоспермі, периспермі насіння, у зародках багатьох рослин запасні білки
зустрічаються у формі алейронових зерен. Алейронові зерна утворюються в процесі визрівання насіння. При цьому
вакуолі в клітинах підсихають, втрачають воду, а білкові речовини, які
містилися до цього в клітинному соку в розчиненому стані, кристалізуються в
алейронове зерно. Одночасно з білками кристалізуються мінеральні солі. Алейронові зерна бувають двох типів: прості й
складні. Просте алейронове зерно (рис. 1.2.2.5) складається з аморфної білкової маси і оболонки, яка її
оточує (бобові, кукурудза, рис).
Рис.1.2.2.5. Запасні
речовини у клітині сім’ядолі насіння квасолі: 1 – крохмальні зерна;
2 – прості алейронові зерна Складне алейронове зерно (рис. 1.2.2.6) має
оболонку і аморфну білкову масу, до складу якої входять кристалоїди (кристалізовані білки) та глобоїди кулястої форми (дрібні кристалічні включення, утворені
мінеральними солями), або тільки кристалоїд різної кутастої форми. Крім того, до аморфної білкової маси алейронових зерен
можуть входити кристали оксалату кальцію (насінини зонтичних, винограду).
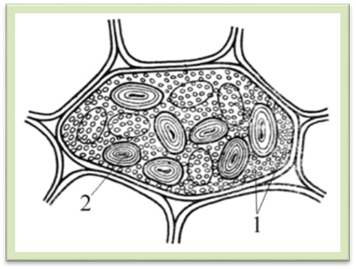
Рис.1.2.2.6. Складні алейронові зерна в ендоспермі
насінини рицини: 1 –
оболонка; 2 – білковий матрикс; 3 – кристалоїди; 4 – глобоїди Під час
проростання насіння білки та глобоїди розчиняються у воді, створюючи поживне середовище
для розвитку зародка рослини. Багато запасних
білків у бобових (чини – 16–28%, волоського горіха – 12–21%, горіха мигдалю – 20–30%, ліщини – 18%, гороху – 23%). Жири і подібні до них речовини. Жири і жирні олії дуже
поширені в тілі рослини і в невеликій кількості напевне зустрічаються в
кожній клітині. Жири – це високоенергетичні сполуки і використовуються в
процесі обміну речовин під час проростання насіння. Вони належать до
найкалорійніших запасних речовин. Термін жир можна використати для
опису не тільки жирів, як ефірів жирних кислот і гліцерину, а й подібних до
них речовин, разом з якими вони об'єднуються в групу ліпідів. Жирні олії
слід розглядати як рідкі жири. До жирів
подібні віск, суберин і кутин, які часто
зустрічаються або в самій клітинній оболонці, або на її поверхні, виконуючи
захисну функцію. Близькі до жирів також фосфатиди і стеарини. Запасні жири розташовані в насінні,
спорах, зародках, в меристематичних клітинах, а інколи і в диференційованих
тканинах вегетативного тіла рослини. Вони зустрічаються в твердому (кокосове масло,
масло бобів какао) (рис.
1.2.2.7) або частіше – рідкому стані.
Рис.1.2.2.7. Тверді рослинні жири: 1 – кокосове
масло; 2 –- какао-масло У рідкому стані запасні жири мають
вигляд краплин різного розміру, розсіяних в цитоплазмі або зібраних в досить
великі маси. Жирові речовини виникають безпосередньо в цитоплазмі, а також
утворюються в лейкопластах (олеопластах). Багато жирів в плодах і насінні: мигдалю – 40–60%, абрикоса – 40%, ліщини – 57–71%, волоського
горіха – 57–74%, маку – 44%.
Багато рослинної олії (рис. 1.2.2.8) у соняшнику, льону, коноплі, ріпаку, рижію, олійної
пальми тощо.
Рис.1.2.2.8. Рослинні олії: 1 – оливкова; 2 – лляна; 3 – рицинова;
4 – кукурудзяна; 5 – соняшникова; 6 – ріпакова Дуже поширені в рослинах ефірні олії – клас дуже
летких ароматичних речовин. У деяких рослин, наприклад у хвойних, вони
зустрічаються в усіх тканинах; в інших рослин ці речовини містяться лише в
пелюстках (троянда), пелюстках
і шкірці плодів (апельсин), корі і
листках (кориця) або в
плодах (мускатника). Біологічне значення ефірних олій полягає в тому, що
своїм запахом вони приваблюють комах-запилювачів або відлякують тварин,
оберігаючи рослину від поїдання. Ефірні олії використовують у парфюмерно-косметичній
промисловості, а також у медицині. Кристали. Мінеральні
речовини у рослин відкладаються головним чином у вигляді солей кальцію і оксидів
кремнію. З солей кальцію найпоширенішою є оксалат, який зустрічається в рослинах у
вигляді моно- і тригідратних солей, утворюючи кристали різного типу (рис.
1.2.2.9). Це – поодинокі кристали, кристалічний пісок (численні дрібні
кристали в одній клітині), друзи, рафіди. Видовжені кристали називають стилоїдами. Друзи – це кристали щавлевокислого кальцію. Вони мають вигляд
зірчастих зростків кристалів різної конфігурації. Рафіди – це пачки голчастих кристалів, що виникають від з'єднання
простих голчастих кристалів.
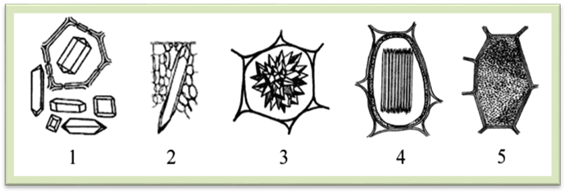
Рис.1.2.2.9. Кристали оксалату кальцію: 1 – поодинокі кристали; 2 – стилоїд; 3 – друзи;
4 – рафіди; 5 – кристалічний пісок Кристали оксалату кальцію звичайно
зустрічаються у вакуолях, але інколи вони утворюються і в цитоплазмі.
Розташовані ці кристали в звичайних клітинах та в спеціалізованих – ідіобластах, які помітно
відрізняються від інших клітин тієї самої тканини за формою і структурою.
Кристали можуть відкладатися в клітинній оболонці. За розмірами кристали бувають дрібні,
менших розмірів, ніж клітини, в яких вони розташовані, або можуть заповнювати
повністю клітину і навіть деформувати її. Рафіди часто зустрічаються в дуже
великих клітинах, які з віком відмирають і заповнені здатним до набрякання
слизом. Під час набрякання слизу оболонка клітини лопається і рафіди
викидаються назовні. Кристали оксалату кальцію відкладаються рівномірно в
усіх клітинах тканини або в певних зонах (наприклад, у винограду вони є в
клітинах, розташованих по краях флоемних променів). Карбонат
кальцію рідко зустрічається у вигляді чітко оформлених
кристалів. Найбільш відомими утвореннями, що включають карбонат кальцію, є
цистоліти (рис. 1.2.2.10) – це
є вирости клітинної оболонки, насичені цією речовиною. Цистоліти зустрічаються в основній
паренхімі, в епідермісі.
Рис.1.2.2.10. Цистоліт в клітині епідермісу листка фікуса Кремнезем
відкладається, головним чином, в клітинах оболонок, але відкладання його
можна спостерігати і в порожнині клітини. Найбільш відомою групою рослин, де
зустрічається кремнезем в аморфному стані як в клітинній оболонці, так і в
порожнині клітини, є родина тонконогових (злакових). Більшість з кристалів є баластними
для рослини, тому рослини скидають листки і гілки, і цим самим регулюють
процес транспірації, а також звільняється від надлишку солей. Вважають, що
кристали можуть виконувати стабілізуючу роль в підтримці осмотичного тиску в
середині клітини. Клітинна
оболонка. У рослинних організмів оболонка целюлозна (рис.
1.2.3.1).
Рис.1.2.3.1. Целюлозні волокна в окремих шарах клітинної оболонки
(мікрофотографія) Наявність міцної оболонки –
характерна особливість рослинної клітини, що відрізняє її від клітини тварин.
Оболонка надає клітині певної форми, створює міцну опору для рослинних тканин
і органів, захищає живий вміст клітини. Як правило, оболонка рослинних клітин
безбарвна, прозора, легко пропускає сонячні промені та воду з розчиненими
низькомолекулярними речовинами. Під час відмирання протопласта
клітини оболонки часто зберігаються і клітини, хоч і мертві, продовжують
виконувати різноманітні функції. Структура клітинної оболонки. Нитчасті
молекули целюлози (С6НІ0О5)n (рис.
1.2.3.2) розташовуються паралельно одна одній і групуються по кілька десятків
таким чином, що виникають тримірні грати, характерні для кристалів. Це так
звані міцели – найдрібніші частки оболонки. Кожна міцела містить до
100 целюлозних молекул. Міцели групуються в мікрофібрили, а останні у фібрили
– волокна або пластинки 0,4-0,5 мкм завтовшки, які видно в світловому
мікроскопі.
Рис.1.2.3.2. Будова целюлозного волокна клітинної оболонки рослин Мікрофібрили целюлози еластичні і
дуже міцні на розрив. Ці властивості зумовлюють широке застосування целюлози
і її продуктів у різних галузях промисловості. Між мікрофібрилами знаходиться
рідкий матрикс, до складу якого входять вода, пектинові речовини і геміцелюлози.
Речовини матриксу визначають такі властивості клітинної оболонки, як
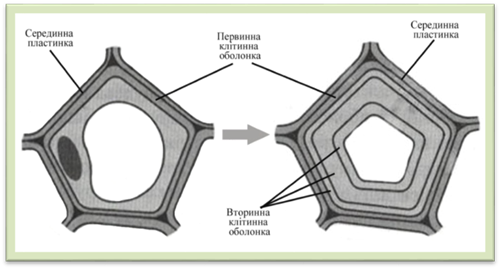
набрякання, високу проникність для води і наявних в ній молекул та йонів. Ріст клітинної оболонки. У процесі
поділу клітини утворені дочірні клітини формують первинну оболонку
Рис.1.2.3.3. Паренхімна клітина з первинною клітинною оболонкою Молода клітина росте доти, доки
розміри її не перестануть змінюватися. Ріст клітини супроводжується
збільшенням її об'єму. При цьому відбувається поверхневий ріст первинної
оболонки внаслідок вклинення нових міцел целюлози в клітинну оболонку між
більш старими, що сприяє її розтягуванню. Цей ріст дістав назву інтусусцепції.
У первинних оболонках паренхімних клітин мікрофібрили розташовані безладно. У
первинних оболонках прозенхімних клітин мікрофібрили розташовані або
паралельно осі клітини (волокниста текстура), або під певним кутом (спіральна
текстура). Оболонки сусідніх клітин ніби
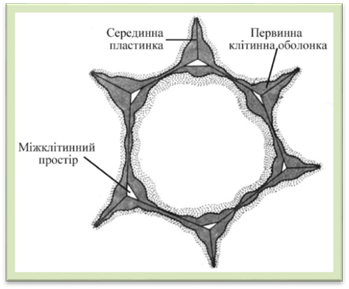
зцементовані між собою міжклітинною речовиною, що утворює так звану серединну
пластинку. Серединна пластинка –
це аморфний міжклітинний шар між первинними оболонками сусідніх клітин. Вона
утворюється першою під час поділу клітин. Складається з пектинових полісахаридів, які за
хімічною природою є поліуроновими кислотами. З клітинної оболонки вони
екстрагуються водою під час тривалого кип'ятіння. У процесі росту оболонки орієнтація
мікрофібрил може змінюватися, що зумовлює шаруватість оболонки. Коли ріст
клітини закінчується, клітинна оболонка може ще потовщуватися. Ріст в товщину
найбільш характерний для вторинних
оболонок клітин (рис. 1.2.3.4) механічних тканин, провідних тощо. Він
здійснюється шляхом послідовного, шар за шаром, відкладання нових міцел
целюлози в напрямку порожнини клітини і призводить до зменшення порожнини. Такий ріст клітинної оболонки
називають апозицією.
Рис.1.2.3.4. Формування вторинної клітинної оболонки Скелетна речовина вторинної оболонки
– целюлоза. Пектинових речовин, геміцелюлоз і води в ній
значно менше, ніж в первинній оболонці. Товщина вторинної оболонки залежить
від спеціалізації клітин і коливається в межах від 1 до 10 мкм. Досить часто
останню частину первинної оболонки, що з'явилась пізніше, особливо, якщо ця
оболонка помітно потовщена, називають вторинною, а самий внутрішній шар
вторинної оболонки розглядають як третинну оболонку. Пори. Порою називають будь-яке непотовщене
місце оболонки у вигляді заглиблення. Самі пори містять найтонші отвори, де
проходять плазмодесми, які з'єднують протопласти сусідніх клітин. Виникають
пори внаслідок нерівномірного потовщення як первинної, так і вторинної
клітинних оболонок. Пори первинних оболонок носять назву первинних порових
полів. У первинних порових полях первинна оболонка відносно тонка, але
неперервна по всій площі порового поля. На цих ділянках оболонки
спостерігається скупчення плазмодесм (видно лише в електронний мікроскоп).
Первинні порові поля характерні для оболонок меристематичних клітин і тих
клітин, які не утворюють вторинних оболонок. У
вторинній оболонці утворюються пори, особливістю яких є те, що на ділянці
первинної оболонки, де розташована пора, вторинна оболонка не відкладається. Пори
вторинної оболонки можуть виникати безпосередньо над первинними поровими
полями, причому кількість їх може варіювати над кожним таким полем від однієї
до кількох. Пори можуть виникати і на тих ділянках первинної оболонки, де
первинних порових полів немає і, навпаки, первинні порові поля можуть бути
повністю перекриті шарами вторинної оболонки. Таким чином, абсолютного
взаємозв'язку між первинними поровими полями і порами немає. Порі,
яка виникла в оболонці будь-якої клітини, звичайно відповідає пора в оболонці
сусідньої клітини, тобто вони утворюються одна проти одної. Таку парну
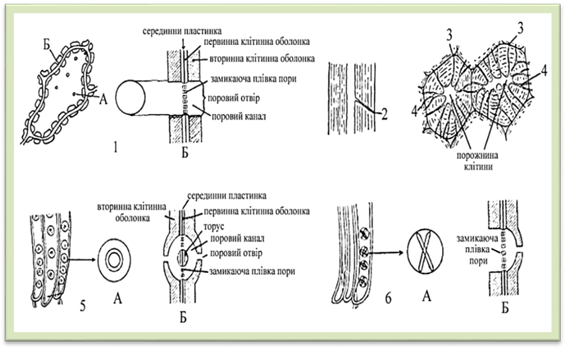
структуру називають парою пор. Замикаюча плівка є загальною для обох
пор і складається із двох первинних оболонок і шару міжклітинної речовини. Пори
бувають прості, облямовані й напівоблямовані (рис. 1.2.3.5). Прості
пори мають циліндричний поровий
канал у межах вторинної оболонки. Вони характерні для основних і твірних
тканин. Напівоблямовані пори
виникають у провідних елементів, що прилягають до основних тканин. З боку
основної тканини формуються прості пори, а з боку провідних – облямовані. Облямовані пори властиві провідним
тканинам; над порами з обох боків утворюються облямівки, які звисають над
первинною оболонкою. Отвір у порожнину клітини називають поровим каналом, порожнину пори – поровою камерою. Первинна оболонка
посередині потовщується, утворює торус, підвішений на еластичній первинній
оболонці, який перекриває отвір облямівки. При великому тиску води він
впирається в облямівки і таким чином захищає паренхімні клітини від їх
руйнування. Вода в горизонтальному напрямі просочується крізь отвори, які є в
торусі і первинній оболонці.
Рис.1.2.3.5. Види та будова пор (А – вид з поверхні, Б – у розрізі): 1 – пряма пора; 2 –
коса пора; 3 – щілиноподібна пора; 4 – розгалужена пора; 5– облямована пора; 6 –
напівоблямована пора Розташовуючись
по-різному, облямовані пори можуть створювати в клітинній оболонці певні
візерунки, які мають спеціальні назви. Розрізняють три основних типи
розташування облямованих пор: драбинчасте,
супротивне і чергове.
Якщо пори мають видовжену або щілиноподібну форму і розташовані одна над
одною – це драбинчаста поровість;
пори, які утворюють короткі горизонтальні ряди, називають супротивною
поровістю; якщо пори
розташовані діагональними рядами, то таку поровість називають черговою. Під дією ферментів у стінках клітин, які
забезпечують функцію руху речовин, утворюються великі отвори, які називають перфораціями. Плазмодесми. Плазмодесми
характерні лише для рослинних клітин. Це дуже тонкі цитоплазматичні нитки Між трубкою ендоплазматичної сітки,
завдяки якій зберігається безперервність ретикулума обох клітин, і
плазмалемою знаходиться гіалоплазма, також безперервна для обох клітин. Коли
клітина перебуває в стані спокою (в насінні, в органах багаторічних рослин),
то цитоплазма відходить від оболонок внаслідок зневоднення вакуоль,
плазмодесми частково або повністю руйнуються і активний зв'язок між клітинами
тимчасово переривається. У кожній клітині міститься до 12
тисяч плазмодесм, які пронизують тонкі ділянки клітинної оболонки – пори.
Плазмодесми, як і пори, і перфорації, служать для транспортування речовин з
клітини в клітину, зв'язку між клітинами.
Рис.1.2.3.6.
Плазмодесми 1.2.4. Хімічні
видозміни клітинної оболонки У
процесі життєдіяльності клітини відбуваються хімічні видозміни целюлозної
клітинної оболонки. Здерев’яніння – це видозміна, яка
характеризується просоченням клітинної стінки лігніном
(рис.1.2.4.1.). Він відкладається в кінці
періоду росту оболонки. Лігнін відкладається нерівномірно: швидше дерев’яніє первинна
оболонка, а потім вторинна і майже не інкрустується третинна. Лігнін заповнює
проміжки між фібрилами клітковини. Відкладання лігніну спричиняє
різке підвищення твердості, міцності оболонки, зникнення її еластичності,
зниження її проникності для води та спричиняє відмирання протопласту.
Рис.1.2.4.1.
Клітинні стінки, просякнуті лігніном Здерев’янілі
клітинні оболонки є стійкими проти руйнування їх мікроорганізмами і грибами. Здерев’яніння забезпечує
міцність стовбурів і гілок дерев. Деревина хвойних та листяних порід містить
до 50% целюлози і 20–30% лігніну. Дерев’яніють і клітинні стінки
багатьох трав, особливо під кінець вегетації. Тому так
важливо своєчасно скосити траву. У рослинному світі може відбуватись роздерев’яніння
клітинної оболонки. Відмічено роздерев’яніння кам’янистих клітин, деревини
плодових дерев, уражених морозами. За сприятливих умов такі пошкоджені
тканини набували здатності до регенерації, особливо клітини з целюлозними
оболонками. Здерев’янілі стінки деяких клітин у плодах груші та айви під час їх
достигання стають м’якими, бо лігнін під впливом ферментів руйнується. Щоб
добути целюлозну масу, з якої виготовляють папір, картон та штучне волокно,
роблять штучне роздерев’яніння деревної маси. Скорковіння – це
видозміна, під час якої зсередини на первинній клітинній оболонці
нашаровується жироподібна речовина – суберин. Клітинні
оболонки стають непроникними для води, газів, спор, шкідників. Протопласт
клітини відмирає і вона заповнюється повітрям. Такі клітини утворюють
відмерлу ізолюючу тканину. За рахунок скорковіння поверхневих шарів пагона
формується корок, який є надійним захистом рослин від температурних коливань
та ураження грибами і шкідниками. Дуб корковий утворює
корок товщиною у кілька сантиметрів і у зв’язку з цим має промислове
використання. Кутинізація – це
видозміна клітинної оболонки, верхні стінки якої покриваються жироподібною
речовиною – кутином, що
відкладається у вигляді суцільної плівочки (кутикули), товщина якої може
досягати 1 см і більше. Кутикула не змочується, непроникна для води, діаспор,
шкідників, надійно захищає листок і пагін від несприятливих температурних
коливань, транспірації. У той же час кутикула не перешкоджає проникненню
сонячних променів і здійсненню фотосинтезу. На зовнішній поверхні оболонки
клітини часто відкладається віск, утворюючи восковий наліт. Він, як і кутикула,
виконує захисну функцію, зменшує випаровування води з поверхні органів. Мінералізація –
відкладання в клітинних стінках солей кальцію та кремнезему. Ці речовини
надають оболонці твердості і крихкості. Збільшення мінералізації клітинних
оболонок хвощів, злаків, осоки відбувається протягом вегетації.
Через це під кінець літа багато з цих рослин настільки просочуються мінеральними
солями, що під час поїдання тваринами викликають кровотечі
від поранення кишечника. Ослизнення –
перетворення целюлози і пектину на слиз і подібні до
них камеді, тобто на
полімерні вуглеводи, які мають здатність дуже набрякати за наявності води
(наприклад, у насінні льону). Утворення
слизу має важливе пристосувальне значення. Під час проростання насіння слиз
закріплює його на певному місці в ґрунті, легко вбирає і утримує воду,
захищає насіння від висихання. Ослизнення клітинних стінок кореневих волосків
забезпечує міцне склеювання їх з частинками ґрунту. Патологічним явищем є виділення слизу
і камеді (рис.1.2.4.2.) внаслідок пошкодження або хворобливого стану рослини
(наприклад, клей у вишні). Ослизнення
у вигляді напливів вишневого клею називають гумозом.
Рис.1.2.4.2.
Виділення камеді Процес
роз'єднання клітин внаслідок руйнування міжклітинної речовини має назву
мацерації. Мацерація буває природна і штучна.
Природна мацерація (рис.1.2.5.1.) відбувається у перестиглих плодах кавуна, дині, груші, сливи та ін. При цьому пектинові речовини серединної пластинки переводяться
в розчинний стан за допомогою ферменту пектинази і потім
вимиваються водою. Природна мацерація відбувається в черешках листків перед
листопадом, в плодоніжках і пелюстках квіток перед їх опаданням. В основі
замочування льону і конопель лежить
мацерація, яку здійснюють мікроорганізми. Штучну
мацерацію проводять під час промислового виділення волокон з прядивних
культур.
Рис.1.2.5.1. Природна мацерація: 1 – у плодах кавуна; 2
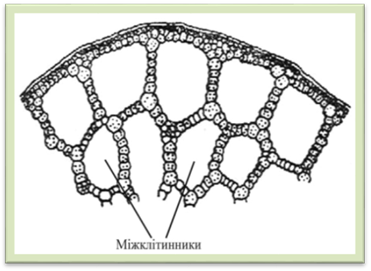
– у пелюстках квіток лотоса 1.2.6. Утворення міжклітинників Під час переходу клітин з
ембріонального до зрілого стану дуже часто спостерігається часткова мацерація
клітин, за якої серединна пластинка руйнується не по всій поверхні, а лише
частково, частіше по кутах клітин. Внаслідок тургорного тиску
сусідні клітини в цих місцях заокруглюються, в результаті чого утворюються
міжклітинники. Спочатку на поперечному розрізі вони мають форму трикутників
або чотирикутників. З ростом оболонок сусідніх клітин міжклітинники
збільшуються і зливаються, утворюючи пов'язану між собою розгалужену сітку,
яка заповнюється парами води і газами. Таким чином, міжклітинники поліпшують газообмін клітин.
Міжклітинники можуть утворюватися і при розчиненні самих клітин. Такі
міжклітинники називають лізигенними. Прикладом їх можуть бути великі
повітроносні міжклітинники у водяних рослин (рис.1.2.6.1), коренях деяких однодольних (Zеа), а також
секреторні вмістища, в які зруйновані клітини виділяють продукт секреції і
частково зберігаються по периферії порожнини.
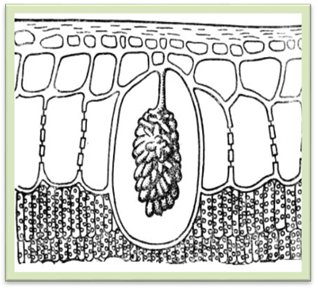
Рис.1.2.6.1. Аеренхіма стебла водяної рослини з великими міжклітинниками Запитання і завдання для самоконтролю 1. Які особливості будови та функцій вакуолей? 2. Які хімічні речовини входять до складу клітинного
соку? Яка їх роль у житті рослин? 3. Назвіть пігменти клітинного соку. Якого забарвлення
вони надають рослинам? 4. Що таке включення? Які речовини належать до
включень? У якому вигляді вони знаходяться у клітині? 5. Які є типи крохмальних зерен? 6. Чим складне алейронове зерно відрізняється від
простого? 7. Які є типи кристалів оксалату кальцію? 8. Яка будова целюлозного волокна клітинної оболонки
рослин? 9. Як відбувається ріст клітинної оболонки рослин? 10. Які є типи пор? 11. Яку роль виконують плазмодесми? 12. Назвіть видозміни клітинної оболонки. Завдяки яким
речовинам здійснюються ці видозміни? 13. Що таке мацерація? Які є види мацерації? 14. Як
утворюються міжклітинники та яка їх роль? |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||