|
|
Ботаніка Електронний посібник |
||
|
|
ЦИТОЛОГІЯ |
||
|
1.1. Протопласт клітини. Поділ ядра і клітини 1.1.2. Методи дослідження клітини. 1.1.3. Різноманітність рослинних клітин. Форма
та розміри клітин. Паренхімні та прозенхімні
клітини. 1.1.4. Протопласт і його компоненти. 1.1.5. Цитоплазма, її фізичні властивості,
хімічний склад і функції. 1.1.6. Мембранна організація цитоплазми. Плазмалема, тонопласт. 1.1.8. Фізіологічно активні речовини цитоплазми. 1.1.9. Ядро, його структура, роль. 1.1.10. Поділ ядра і клітини як основа процесів
розмноження і розвитку рослин. 1.1.11. Поняття про мітотичний цикл. Інтерфаза. 1.1.15. Порушення процесу мейозу. Поліплоїдія. Будь-який рослинний організм складається з клітин (рис 1.1.1.1).
У нижчих рослин клітина може становити собою цілий одноклітинний організм
(наприклад, у хламідомонади, хлорели), або бути складовою
частиною колонії (наприклад, у вольвокса),
або елементом багатоклітинного організму. Для вищих рослин характерні висока
спеціалізація клітин, об’єднання їх у тканини, з яких формуються органи.
Клітини в організмі зв'язані між собою спільним обміном речовин, походженням
і виконанням певних функцій.
Рис.
1.1.1.1. Рослини –
клітинні організми: 1
– хламідомонада; 2 – вольвокс; 3 – улотрикс: 4 – валіснерія і клітинна будова листка Клітина – це основна структурна і функціональна одиниця живої
матерії, елементарна біологічна система, що здатна до самовідтворення. Наука про мікроскопічну і субмікроскопічну структуру
клітини та її життєдіяльність називається цитологія. Уперше клітину
побачив англійський природодослідник Роберт Гук (рис 1.1.1.2), вивчаючи за допомогою
сконструйованого ним світлового мікроскопа зріз корку (покривної тканини)
бузини. Він встановив, що корок складається з дрібних комірок, відмежованих
оболонками. Свої спостереження Р. Гук описав у праці «Мікрографія»,
яку було видано у 1665 році. У цій роботі він уперше назвав ці комірки
«клітинами». Оскільки корок складається з мертвих клітин, які мають лише
стінки, виникло помилкове уявлення про те, що
стінки виконують основні життєві функції клітини. Вмісту клітин надавали
другорядного значення, вважаючи його «живильним соком» або «рослинним
слизом». І лише у ХІХ ст. дослідники змінили свою думку про роль вмісту
клітини. На цей час уже були відомі крохмальні
зерна, кристали, хлоропласти та інші частини клітини. Вдосконалювалась
мікроскопічна техніка, накопичувались нові експериментальні дані про
внутрішній вміст клітини.
Рис. 1.1.1.2. Відкриття клітини: 1 – Роберт Гук, 2 – мікроскоп Гука, 3 – клітини корка У процесі
подальшого вивчення клітини дослідники почали звертати більше уваги не на
оболонку, а на внутрішній вміст, якому в 1880 р. німецький вчений Й. Ганштейн дав назву «протопласт». У 1831 р. англійський
ботанік Р. Броун
Рис. 1.1.1.3. Вчені-ботаніки: 1 – Р. Броун, 2 – Г. Моль В 1838 р.
німецький ботанік М. Шлейден довів,
що клітина та її органели є складовими рослинного організму, а в 1839 р. його
співвітчизник зоолог Т. Шванн (рис
1.1.1.4), спираючись на праці
М. Шлейдена, сформулював положення клітинної теорії, основні з яких: усі живі організми складаються з клітин;
клітини тварин і рослин подібні за будовою. Клітинна теорія будови
організмів мала великий вплив на розвиток не тільки ботаніки, а й всього
природознавства. Вона спрямовувала зусилля вчених біологів на детальніше
вивчення вмісту клітини. Клітинною теорією було доведено єдність походження,
будови та еволюції рослин і тварин.
Рис. 1.1.1.4. Вчені – основоположники клітинної
теорії: 1 – М. Шлейден, 2 – Т. Шванн У другій половині ХІХ
ст. було зроблено нові відкриття, які збагатили клітинну теорію. Російський
вчений
Рис. 1.1.1.5. Вчені, які збагатили клітинну теорію: 1 – П.Ф. Горянінов, 2 – І.Д. Чистяков, 3 – Е. Страсбургер, 4 – І.М. Горожанкін Сучасна клітинна теорія наповнилась новими
відомостями. Суть її полягає в тому, що: ·
клітина
– елементарна структурна одиниця живих організмів; ·
клітини
різних організмів подібні за походженням, будовою, хімічним складом;
основними процесами життєдіяльності; ·
кожна
нова клітина утворюється внаслідок поділу материнської; ·
багатоклітинні
рослинні організми – це складні комплекси клітин, поєднані між собою у
цілісні системи тканин, з яких складаються органи. На кінець ХІХ ст.
цитологія остаточно сформувалась у самостійну науку. Завдяки світловому
мікроскопу було вивчено основні компоненти клітин, накопичувались відомості
про їхні функції. Подальший прогрес цитології пов’язаний з винайденням
електронного мікроскопа. В біології його почали використовувати в середині ХХ
ст. Під електронним мікроскопом було розглянуто важливі деталі структури
відомих компонентів клітини, а також відкрито нові. Нині цитологія є однією з біологічних наук, що
розвивається дуже інтенсивно. Перед нею поставлене
завдання вивчити деталі структури компонентів клітини, її спадковості,
захисних пристосувань (імунітету); змін під дією факторів навколишнього
середовища тощо. Розвиток нових галузей цитології – фізіології, біохімії, фізикохімії
компонентів клітини – дасть змогу розгадати одну з таємниць природи – суті
самого життя. 1.1.2. Методи дослідження клітини Методи, що застосовують для вивчення клітини, дуже
різноманітні. Основний з них мікроскопічний.
Велику роль відіграє світловий
мікроскоп, сучасні моделі якого дають збільшення до 2–3 тис. разів. За
допомогою світлового мікроскопа спостерігають тонкі зрізи в прохідному і
відбивному світлі. Метод світлової мікроскопії ґрунтується на тому, що через
прозорий чи напівпрозорий об’єкт дослідження проходять промені світла, які
згодом потрапляють до системи лінз об’єктива та окуляра. Ці лінзи збільшують
об’єкт дослідження, при цьому кратність збільшення можна визначити як добуток
збільшень об’єктива та окуляра. Однак внаслідок природи самого світла
можливості оптичного мікроскопа обмежені, часточки менші як 0,2 мкм (1 мкм (мікрометр) = 0,001
мм) розглянути під таким мікроскопом неможливо. Клітинні структури найдрібніших розмірів були відкриті
і вивчені за допомогою електронного
мікроскопа, винайденого в першій половині ХХ сторіччя. В даний час широко використовуються трансмісійні (просвічуюючі)
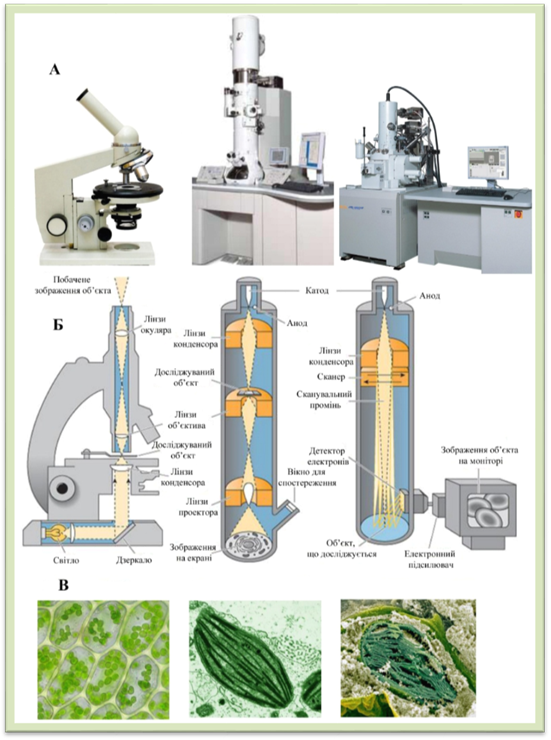
електронні мікроскопи і сканувальні (растрові) електронні мікроскопи (рис 1.1.2.1). Електронний мікроскоп
дає збільшення об’єктів дослідження до 500 тис. разів і більше. В ньому
замість променів світла застосовують потік електронів, які рухаються в
магнітному полі з величезною швидкістю. Роль лінз при цьому виконують
електромагніти, здатні змінювати напрямок руху електронів, збирати їх у пучок
(фокусувати) й спрямовувати його на об’єкт дослідження. У трансмісійному
електронному мікроскопі електрони, пройшовши через досліджуваний об’єкт,
потрапляють на люмінесцентний екран, спричиняючи його нерівномірне свічення,
або на особливий фотоматеріал, за допомогою якого зображення можна
фотографувати. Зрізи, що вивчаються, повинні бути не більше як 0,05 мкм завтовшки і мати спеціальне забарвлення. За допомогою
електронного мікроскопа можна розглянути структуру розміром 1,5 нм (1 нм
(нанометр) = 0,001 мкм). Поверхні клітин, окремих органел тощо можна вивчати за
допомогою сканувальних електронних мікроскопів, які здатні
створювати тривимірне зображення. В них потоки електронів не проходить крізь
об’єкт дослідження, а відбивається від його поверхні, на який напилено метал.
Рис.
1.1.2.1. (зліва направо): А. Загальний вигляд мікроскопів: світлового, трансмісійного електронного, скануючого електронного. Б. Схеми оптичних шляхів мікроскопів: світлового, трансмісійного електронного, скануючого електронного. В. Фотографії хлоропластів
зроблені
за допомогою світлового, трансмісійного електронного, скануючого електронного мікроскопів Методом культури тканин вивчають структуру та
життєдіяльність живих клітин поза організмом. При цьому живі клітини
утримують та розмножують на штучних поживних середовищах. Змінюючи компоненти
поживного середовища, можна спостерігати, як ті чи інші сполуки впливатимуть
на ріст і розмноження клітин, інші їхні властивості. Культури клітин
використовують у медицині, ветеринарії та службі захисту рослин для перевірки
впливу різноманітних хімічних препаратів, вірусів, одноклітинних організмів,
отримання біологічно активних речовин. На основі цього методу розроблено
клітинні технології, за допомогою яких конструюються організми із заданими
наперед властивостями. Так, із кількох рослинних клітин удається виростити
цілу рослину. Метод мічених
атомів, або авторадіографія, дає змогу з’ясувати місце та перебіг
певних фізико-хімічних явищ у клітині. Для цього до клітини вводять речовину,
в якій один з атомів певного елемента (Карбону, Фосфору тощо) заміщений його
радіоактивним ізотопом. За допомогою особливих приладів, здатних виявляти
ізотопи, можна прослідкувати за міграцією цих речовин у клітині, їхніми
перетвореннями, виявити місце та характер тих чи інших біохімічних процесів. Цитохімічний метод
пов'язаний з дослідженням хімічного складу клітин. Цим методом можна виявити
наявність і визначити кількість різних речовин у клітині – білків , жирів,
вуглеводів, нуклеїнових кислот, гормонів, вітамінів, тощо. За допомогою методу
центрифугування можна
розділити компоненти клітини, що мають різну густину з метою їх вивчення. При
цьому клітини попередньо подрібнюють і в особливих пробірках поміщають у
центрифугу – прилад, здатний розвивати швидкі оберти (рис. 1.1.2.3). Оскільки
різні клітинні структури мають неоднакову щільність, при дуже швидких обертах
центрифуги вони осідатимуть шарами: щільні органели – швидше і тому опиняються
знизу, а менш щільні – зверху. Ці шари розділяють і вивчають окремо
мікроскопією та іншими методами.
Рис.
1.1.2.3. Центрифуга лабораторна Методом мікроскопічної хірургії можна
добути з клітини певні компоненти (ядро, мітохондрії тощо), здійснити перенос
окремих органел з однієї клітини до іншої. За допомогою мікроманіпулятора
(рис. 1.1.2.4) клітини
розрізають, витягують з них частини, вводять речовини (мікроін’єкції).
Мікроманіпулятор поєднують з мікроскопом, в який спостерігають за ходом
операції. Завдяки мікрохірургії було виявлено, що ядро визначає форму клітини
і характер її функціонування.
Рис.
1.1.2.4. Мікроманіпулятор 1.1.3. Різноманітність
рослинних клітин. Форма та розміри клітин. Паренхімні та прозенхімні
клітини Форма та розміри клітин дуже різноманітні і залежать
від місця їх розташування та функції, яку вони виконують. Так, клітини конуса наростання ізодіаметричні,
але спеціалізуючись на виконанні фізіологічних функцій, вони змінюють свою
форму і будову. Клітини зовнішнього шару, що дають початок епідермісу,
призматичні або плоскі. Клітини центральної частини конуса наростання
багатокутні, подовжені, вони дають початок провідним, основним та іншим
тканинам. Клітини епідермісу плоскі із звивистими оболонками. Клітини, які
проводять пластичні речовини, трубчасті з тонкостінною целюлозною оболонкою;
клітини механічних тканин багатокутні з потовщеними оболонками та відмерлим
внутрішнім вмістом. Зірчасту форму мають клітини аеренхіми. Клітини, які
виконують функцію запасу поживних речовин мають округлу, овальну, призматичну
форму. Найчастіше клітини, які зазнають взаємного тиску мають
форму багатокутників. Форма клітин, що ростуть вільно, може бути
кулеподібною, зірчастою, циліндричною тощо. Всі клітини за формою поділяються на дві групи: паренхімні і прозенхімні (рис. 1.1.3.1). Паренхімні клітини, в
основному, мають більш-менш однакову довжину і ширину, або їх ширина
становить 2/3 довжини. За формою вони бувають овальні, округлі, багатокутні.
Терміном «паренхіма» називають тканину, що складається з тонкостінних живих
клітин, які пов’язані з різними процесами життєдіяльності клітин, такими як
фотосинтез, дихання, запас поживних речовин. Середній розмір клітин – 10–100
мікронів (1мк = 0,001 мм) за винятком клітин бактерій (0,5–5 мк), які можна побачити лише при максимальному збільшенні
світлового мікроскопа. Паренхімні клітини, в яких відкладаються запаси води
та поживних речовин досягають у довжину кількох міліметрів, їх видно
неозброєним оком, наприклад клітини кавуна, помідора, лимона, бульб картоплі. Прозенхімні клітини – це клітини витягнуті, у
яких довжина перевищує ширину у багато разів. До них належать волокна механічних
тканин, камбіальні клітини, клітини коленхіми, ситоподібних трубок та ін.
Особливо довгі прозенхімні клітини стебла льону – 40 мм, волосків бавовнику – 60 мм, кропиви – 80 мм, рамі – 200 мм. Діаметр цих
гігантських клітин набагато менший.
Рис.
1.1.3.1. Форми клітин листка моху мніума (фотографія зроблена за
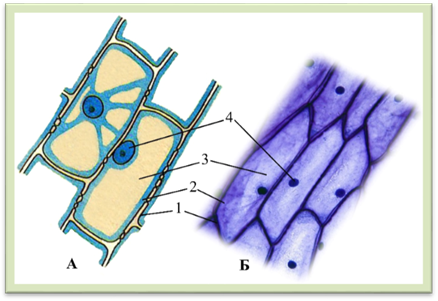
допомогою світловогоого мікроскопа): 1 – паренхімні, 2 – прозенхімні 1.1.4. Протопласт і його компоненти Незважаючи на величезну різноманітність, всім клітинам
притаманна спільність будови. Розглядаючи дорослу клітину за допомогою світлового
мікроскопа, можна побачити такі компоненти: клітинну стінку, цитоплазму, вакуолі, ядро (рис. 1.1.4.1).
Рис.
1.1.4.1. Клітини епідермісу соковитої луски цибулі: А – схема будови, Б – фотографія зроблена
за допомогою світлового мікроскопа; 1 – клітинна стінка, 2 – цитоплазма, 3 –
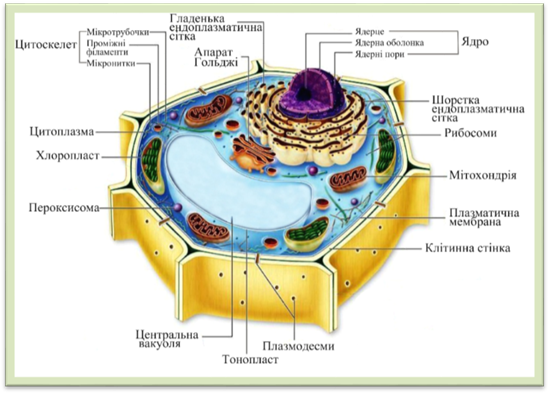
вакуоля, 4 – ядро з ядерцем Дослідження клітини під електронним мікроскопом виявили
значно складнішу будову рослинних клітин
Рис. 1.1.4.2. Схема будови рослинної клітини за даними електронного мікроскопа Розрізняють дві групи компонентів в клітині: перша –
активні або живі частини, їх сукупність називають протопластом (рис. 1.1.4.3). До них
відносять цитоплазматичну мембрану, цитоплазму з органелами і ядро. Друга
група компонентів – неактивні або неживі частини клітини. До них відносять
клітинну стінку, вакуолі, включення. Вони є продуктами життєдіяльності
протопласта, тому їх називають похідними протопласта. Протопласт займає
найбільшу за об'ємом і найважливішу за значенням частину клітини. З
життєдіяльністю протопласта зв'язані всі метаболічні процеси, розмноження,
дихання, реакція на зовнішні подразнення тощо.
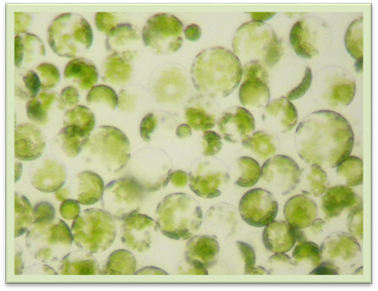
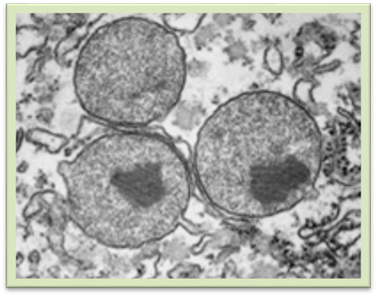
Рис. 1.1.4.3. Протопласти клітин листка петунії 1.1.5. Цитоплазма, її фізичні властивості,
хімічний склад і функції Цитоплазма. Неодмінною
складовою живої рослинної клітини є цитоплазма – безбарвна, зерниста,
більш-менш в’язка рідина, в якій проходять найважливіші життєві процеси –
синтез, дихання, ріст, рух тощо. Цитоплазма існує в процесі безперервного обміну
речовин, в результаті якого відбувається її ріст та розвиток, збільшення маси
й диференціація. Цитоплазма реагує на впив зовнішніх факторів (світла,
температури, сили тяжіння тощо), тобто характеризується подразливістю. За хімічним складом цитоплазма неоднорідна. До її
складу входять амінокислоти, білки, вуглеводи, ліпіди, нуклеїнові кислоти,
вода, неорганічні сполуки, вітаміни, ферменти, фітонциди, органічні кислоти
та ін. У клітині 70–80% води. Вода є природним розчинником органічних
та неорганічних сполук, умовою ферментативної
активності, здійснення фотосинтезу, метаболізму клітини та рослини в цілому.
У цитоплазмі вода знаходиться у вільному та зв’язаному стані. Більше
зв’язаної води міститься у плодах і насінні (соя, пшениця, квасоля, гречка). У молекулярній та структурній організації цитоплазми
важливе місце займають білки (10–20%). До складу всіх білків входять атоми
Карбону, Гідрогену, Оксигену, Нітрогену. Крім того,
до складу багатьох білків входять атоми Сульфуру. Є
білки, до складу яких входять атоми Фосфору, Феруму
та інші. Білки – це
високомолекулярні біополімери, мономерами яких є амінокислоти. До складу
білків входять 20 основних амінокислот. Білки, що складаються лише з
амінокислот, називають простими або
протеїнами. Складні білки або протеїди утворюються внаслідок
сполучення простих білків з вуглеводами, ліпідами, нуклеїновими кислотами. За
природою білки є конституційні та аморфні: Перші витрачаються на
формування клітинних структур, а другі включаються у загальний обмін речовин
або відкладаються про запас. Важливу роль у цитоплазмі відіграють вуглеводи, які
утворюються в процесі фотосинтезу. Вони знаходяться в цитоплазмі у твердому
або рідкому станах. Частина з них відкладається про запас, а основна маса
використовується для побудови структурних частин органел і створення
енергетичного потенціалу клітини. У цитоплазмі міститься 2–3% ліпідів. Взаємодіючи з білками, вони утворюють ліпопротеїдний
комплекс, що має важливе значення у формуванні структурних елементів органел
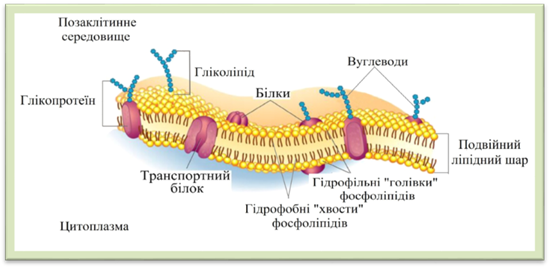
клітини, особливо клітинних мембран різної природи та призначення. 1.1.6. Мембранна організація цитоплазми. Плазмалема, тонопласт Цитоплазма
має мембранну організацію. Її структуру утворюють тонкі (4–10 нм), досить
щільні плівки – біологічні мембрани (рис. 1.1.6.1). Мембрана
складається з ліпідів, білків, вуглеводів. Молекули ліпідів (фосфоліпідів)
розташовані у два шари, так що їхні гідрофільні частинки «голівки», які інтенсивно взаємодіють з водою, обернені до зовнішнього
та внутрішнього боку мембрани, а гідрофобні частинки «хвости», інертні до
води, – спрямовані всередину. Молекули білка розташовані несуцільним шаром на
поверхні ліпідного каркаса з обох його боків. Частина їх заглиблена в ліпідний
шар, а деякі проходять крізь нього наскрізь, утворюючи ділянки, проникні для
води. Вуглеводи входять до складу мембран виключно у вигляді комплексних сполук з молекулами білків (глікопротеїни)
або ліпідів (гліколіпіди). Мембрани утворюють пограничний шар цитоплазми, а також
зовнішню межу її органел і беруть участь в утворенні їхньої внутрішньої
структури. Вони поділяють цитоплазму на ізольовані відсіки, в яких одночасно
і незалежно один від одного можуть проходити біохімічні процеси, часто у протилежному
напрямку (наприклад, синтез і розпад). Однією з основних властивостей біологічних мембран є вибіркова проникність, або напівпроникність,
завдяки чому деякі речовини проникають крізь них погано, інші легко і навіть
у напрямку більшої концентрації. Таким чином, мембрани визначають хімічний
склад цитоплазми. Плазмалема або плазматична мембрана являє собою відносно тонкий зовнішній
шар цитоплазми, відмежований від оболонки клітини. Власне, це мембрана, яка
відмежовує цитоплазму від стінки клітини і звичайно щільно прилягає до неї.
Іноді плазмалема буває хвилястою або утворює
глибокі складки. Вона регулює обмін речовин клітини з навколишнім
середовищем, а також бере участь у синтезі речовин.
Рис. 1.1.6.1. Схема будови плазматичної мембрани Тонопласт, або вакуолярна мембрана, відділяє цитоплазму від вакуолей
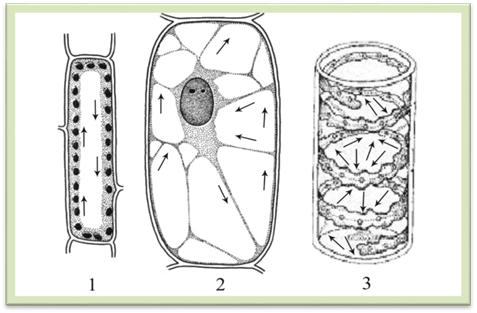
та виконує ті самі функції, що й плазмалема. Решта цитоплазми – мезоплазма – міститься між плазмалемою і тоноплластом. Мезоплазма складається з однорідної безструктурної гіалоплазми і структурних органел. Гіалоплазма – це рідке
неперервне середовище, в яке занурені органели. Властивість гіалоплазми як
складової частини цитоплазми – рух, яким регулюється інтенсивність обміну
речовин у клітині. Він стає енергійнішим під час посиленої її діяльності. Рух
буває круговий – відбувається
уздовж стінки клітини в одному напрямку, якщо в центрі міститься одна
велика вакуоля (листок елодеї); струменистий
або циркулярний – по тяжах, які перетинають центральну вакуолю у різних
напрямках (волоски кропиви, гарбуза); коливальний
– коли дрібні частки, які розташовані у внутрішньому шарі цитоплазми,
плавно рухаються в одному напрямку (наприклад, у водоростей), а деякі
частинки цитоплазми перебувають у стані спокою, тому цей рух найменш
впорядкований (рис. 1.1.6.2). Швидкість руху залежить від температури, інтенсивності
світла, забезпечення киснем та інших факторів. Під час руху гіалоплазма
захоплює з собою органели. Роль гіалоплазми велика: в ній здійснюється
взаємозв’язок між органелами, вона бере участь в обміні речовин, передачі
подразнення тощо. Гіалоплазма є середовищем для взаємодії речовин.
Рис. 1.1.6.2. Типи руху цитоплазми (напрямок руху
показано стрілками): 1 – круговий у клітині листка
елодеї, 2 – струменистий у клітині волоска тичинкової нитки
традесканції, 3 – коливальний у клітині спірогіри 1.1.7. Структурні компоненти
цитоплазми (ендоплазматична сітка, апарат Гольджі, сферосоми, лізосоми, мікротільця, рибосоми, мікротрубочки,
мікрофіламенти, мітохондрії, пластиди), особливості
їх будови і функції Органели найчастіше поділяють на дві групи: ті, що видно
під світловим мікроскопом, і ті, що видно лише під електронним мікроскопом. У
кожній групі розрізняють такі органели: вкриті двома мембранами, вкриті
однією мембраною і немембранні. Ендоплазматична
сітка (ендоплазматичний ретикулум) являє собою
густу, розгалужену по всій цитоплазмі сітку взаємопов’язаних, обмежованих
мембраною ультрамікроскопічних канальців, пухирців, трубочок і цистерн. Мембрани ендоплазматичної сітки утворюють єдину систему
з мембраною ядерної оболонки, в той же час вони не сполучаються ні з плазмалемою, ні з тонопластом.
Мембрани ендоплазматичної сітки сусідніх клітин з’єднуються між собою
трубочками, що проходять по плазмодесмах крізь пори у клітинній оболонці. Розрізняють два різновиди ендоплазматичної сітки – гранулярна (шорстка) і агранулярна
(гладенька) (рис. 1.1.7.1). Гранулярна
ендоплазматична сітка, яка утворена переважно цистернами,
має на поверхні дрібні гранули – рибосоми. Тут відбувається за участі рибосом
біосинтез білка, поліпептидний ланцюг якого переноситься до внутрішнього
простору структур ендоплазматичної сітки. Агранулярна
ендоплазматична сітка складається з розгалужених трубочок,
які відходять від цистерн гранулярної ендоплазматичної сітки, на його
поверхні немає рибосом. У ній відбувається біосинтез вуглеводів
(полісахаридів, зокрема целюлози), а також ліпідів. У молодих і багатих на білки клітин переважає
гранулярна ендоплазматична сітка, а у старин і багатих на вуглеводи і ліпіди
– агранулярна. Ендоплазматична сітка забезпечує внутрішньоклітинний і міжклітинний
транспорт, синтез білків, ліпідів, полісахаридів і перенесення їх у різні
частини клітини, зборку клітинних мембран. У мембранах ендоплазматичної сітки
міститься багато різних ферментних систем, що має важливе значення для обміну
речовин у клітині.
Рис. 1.1.7.1. Ендоплазматична сітка: А – схема будови гранулярної
ендоплазматичної сітки, Б – схема будови агранулярної
ендоплазматичної сітки; 1 – фотографія гранулярної
ендоплазматичної сітки зроблена під електронним мікроскопом, 2 – фотографія агранулярної ендоплазматичної сітки зроблена під
електронним мікроскопом Апарат
(комплекс) Гольджі являє собою систему, що складається з
окремих елементів – диктіосом.
Кожна диктіосома побудована з 5–8 сплющених цистерн
завтовшки 1–2 мкм, обмежованих мембраною, які
лежать паралельно. Між цистернами знаходиться плівка завтовшки 10 нм.
Цистерни можуть мати форму диска з гладкими здутими краями або мати по краю
диска сітку розгалужених трубочок із здуттям на кінцях. Цистерни оточені
численними пухирцями, що відшнуровуються від їх
периферичних ділянок (рис. 1.1.7.2).
Рис. 1.1.7.2. Апарат Гольджі: А – схема будови, Б –
фотографія зроблена за допомогою електронного мікроскопа Апарат Гольджі безпосередньо пов'язаний з
ендоплазматичною сіткою як просторово, так і функціонально. Канальцями
ендоплазматичної сітки до комплексу Гольджі транспортуються продукти
біосинтезу, які потім накопичуються в пухирцях. Ці пухирці відчленовуються від цистерн і надходять у цитоплазму та
вакуолі. Різні сполуки, що містяться у пухирцях, які надходять у цитоплазму
(пектин, геміцелюлози), використовуються рослиною для побудови матрикса первинної клітинної оболонки, а мембрани
пухирців вбудовуються в плазмалему молодих клітин.
В пухирцях диктіосом можуть накопичуватись ефірні
олії, слиз або деякі речовини вторинного синтезу. Ці пухирці, відчленовуючись від цистерн, надходять у вакуолі. Вони
виконують секреторну функцію. Отже, комплекс Гольджі є органелою, в якій нагромаджуються, конденсуються
і пакуються матеріали, що виводяться назовні. Він є одним з кінцевих етапів
клітинного «конвеєра» формування включень. Розвивається органела з елементів
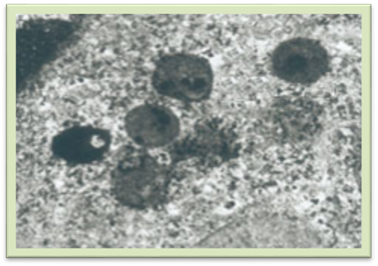
ендоплазматичної сітки. Лізосоми – це пухирці 0,2-0,8мкм діаметром, оточені мембраною (рис. 1.1.7.3). Містять комплекс
гідролітичних ферментів, які розщеплюють нуклеїнові кислоти, білки, вуглеводи,
ліпіди. Продукти розщеплення виходять у цитоплазматичний матрикс,
де й використовуються, або разом з пухирцями надходять до вакуолей.
Лізосоми можуть руйнувати окремі органели або
ділянки цитоплазми (локальний автоліз) у разі потреби оновлення клітин.
Рис.
1.1.7.3. Лізосоми (фотографію зроблено за допомогою електронного
мікроскопа) Сферосоми – округлі тільця
0,2–1,3 мкм діаметром, оточені мембраною. Сферосоми відшнуровуються від
кінців тяжів ендоплазматичної сітки. Сферосоми
синтезують і накопичують рослинні жири. В міру синтезу жирів матрикс в сферосомі
витісняється, мембранна оболонка редукується і від неї залишається лише
зовнішній шар. Тому дозріла сферосома є краплиною
жиру, оточеною тонкою мембраною. Мікротільця – це обов’язкові
органели еукаріотчної рослинної клітини.
Розрізняють кілька видів мікротілець: пероксисоми, пероксисоми листків, гліоксисоми, неспеціалізовані пероксисоми.
Пероксисоми – це малі органели
рослинної клітин. Частіше трапляються на внутрішніх мембранах – кристах мітохондрій, ендоплазматичній сітці, інших
структурах. За походженням – це похідні ендоплазматичної сітки, від якої
відособлюються або залишаються з’єднаними з нею. Пероксисоми
кулястої форми, діаметром 0,15–1,5 мкм (рис. 1.1.7.4). Вони складаються із
строми, або матриксу, оточеного одинарною
мембраною. У деяких клітин матрикс диференційований
на тонкозернисту аморфну периферійну та щільну центральну частину,
представлену кристалоїдом, утвореним волокнистими або трубчастими
структурами. Пероксисоми – це білково-ліпідні
тільця, які містять ферменти окислення амінокислот. При цьому виникає
гідроген пероксиду (перекис водню), який є токсичним для клітин. Роль
детоксикації здійснюється з участю ферменту каталази, який є одним з найбільш
швидкодіючих: він розщеплює гідроген пероксиду до води та кисню, звідки й
походить сама назва пероксисоми.
Рис.
1.1.7.4. Пероксисоми (фотографію зроблено за допомогою електронного
мікроскопа) Пероксисоми листів беруть участь у
фото диханні та фотосинтезі зелених рослин. Субстратом для фото дихання є гліколева кислота, яка синтезується в процесі
фотосинтезу. Отже, пероксисоми – це активні дегратогенні чинники ендогенних і екзогенних молекулярних
сполук клітини. Гліоксисоми – це органели, які трапляються в клітинах ендосперму
насіння, багатого на олію. За їх участю в процесі гліоксилатного
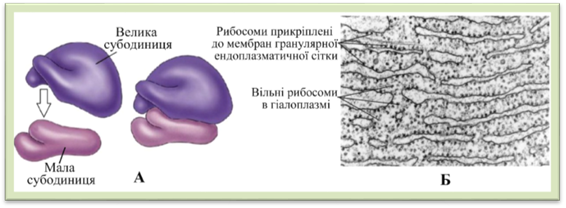
циклу ліпіди трансформуються в жири. Рибосоми – це
сферичні гранули близько 20 нм діаметром, які під електронним мікроскопом
видно у вигляді темних цяточок. Вони виявлені також у мітохондріях і
пластидах, але мають менші розміри. Рибосоми розташовуються вільно в
гіалоплазмі або прикріплені до поверхні мембран гранулярної ендоплазматичної
сітки. Мембранної будови рибосоми не мають. Кожна рибосома складається з двох
субодиниць – великої і малої, побудованих з білка
та рибосомної рибонуклеїнової кислоти (рРНК) (рис. 1.1.7.5). Субодиниці рибосом формуються у ядрі за участю ядерець.
Рис.
1.1.7.5. Рибосоми: А – схема будови, Б – фотографія зроблена за
допомогою електронного мікроскопа Функція рибосом – біосинтез білка. Цей процес відбувається
в рибосомах, розташованих групою і пов’язаних між собою ниткоподібною молекулою інформаційної
(іРНК). Такі групи рибосом називаються полісомами. У полісоми нараховується до
30–40 одинарних рибосом. На одній полісомі
може водночас відбуватись синтез багатьох молекул певного білка. Мікрофіламенти – органели
рослинної клітини. За соєю природою це білкові нитчасті структури діаметром
5–7 нм, що належать до групи скорочувальних білків-актинів
і складаються з великої кількості субодиниць,
згрупованих у спаралізовані стрічкоподібні утворення. Завдяки скорочувальності актину та виникаючого при цьому тертя
одного об інший щільно розміщених цитоплазматичних тяжів, спричиняється рух
цитоплазми. За участю мікрофіламентів у клітині
виникають різні види руху цитоплазми та її органел. Напрям руху спрямовується
системою мікрофіламентів. Ці структури є
обов’язковим компонентом клітин майже всіх еукаріотичних рослин і відіграють
істотне значення у здійсненні метаболічних процесів. Разом із системою мікротрубочок мікрофіламенти
утворюють рухливу сітчасту структуру протопласту, яку називають цитоскелетом клітини (рис. 1.1.7.6). Мікрофіламенти
не довговічні, замість зникаючих старих з’являються нові.
Рис.
1.1.7.6. Схема будови цитоскелета Мікротрубочки. У цитоплазмі
клітин еукаріотичних рослин під електронним мікроскопом чітко виділяються
тонкі порожнисті структури завдовжки 0,5–3,5 мкм і
діаметром 24 нм, які називають мікротрубочками. Подібні
циліндричні структури утворені 13 поздовжніми нитками. Кожна з них
складається з безлічі розміщених сферичних глобулярних субодиниць
білка тубуліну. Сформована мікротрубочка
має діаметр порожнини 19–29 нм і товщину оболонки 5 нм. Мікротрубочки
у клітині утворюють динамічну і недовговічну систему, у живих рухливих клітин
генетично давні елементи чи цілі мікробрубочки
руйнуються і зникають, а замість них зявляються
нові, відновлюючи їх систему і функціональну активність. Мікротрубочки
наростають шляхом приєднання до одного з кінців нових тубулінових
субодиниць. У рослинній
клітині мікротрубочки виконують різноманітні
функції. У молодих ембріональних клітин вони розміщуються з внутрішнього боку
плазмалеми, визначаючи характер формування і
групування целюлозних мікрофібрил, а також їх
послідовне включення у наростаючу клітинну оболонку. За допомогою мікротрубочок пухирці Гольджі переміщуються до
наростаючої клітинної пластинки. Крім того, за їх участю від ендоплазматичної
сітки, з якої відособлюються мікропухирці,
відбувається їх потік до комплексу Гольджі. В клітині переміщуються й більші
за розмірами органели (мітохондрії, лізосоми). Мікротрубочки забезпечують просторове розміщення та
пересування органел до місця фізіологічної активності, розходження хромосом
до полюсів при діленні ядра. Ці структури беруть участь у формуванні
первинної клітинної пластинки між дочірніми клітинами у процесі цитокінезу. Вони є компонентами джгутиків, війок, центріолей, ахроматинових
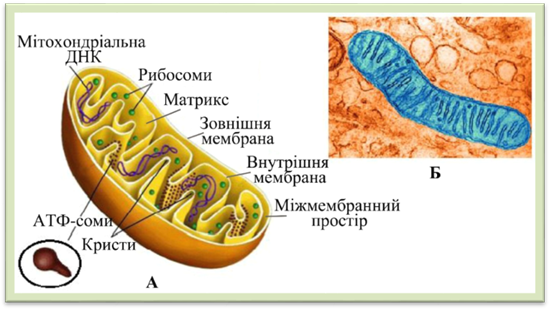
ниток. Мітохондрії являють собою органели округлої, паличкоподібної, ниткоподібної форми. В
одній клітині залежно від типу і стану їх міститься від 50 до 5000. Мітохондрії є двомембранними органелами. Зовнішня мембрана мітохондрії
гладка, а внутрішня утворює вирости у порожнину мітохондрії у вигляді складок
або трубочок, які називають кристами. Завдяки кристам значно
збільшується внутрішня мембранна поверхня мітохондрій. Простір між кристами заповнено рідкою речовиною – матриксом, в якій знаходяться рибосоми, дезоксирибонуклеїнова
кислота (ДНК), інформаційна (іРНК), транспортна (рРНК) рибонуклеїнові кислоти . Поверхня внутрішньої
мембрани вкрита дуже дрібними тільцями – АТФ-сомами,
які мають кулясту головку та ніжку. Саме на них зосереджені ферментні системи
мітохондрій (рис. 1.1.7.7).
Рис.
1.1.7.7. Мітохондрія: А – схема будови, Б – фотографія зроблена за
допомогою електронного мікроскопа Мітохондрії є енергетичними
центрами клітини. В них у процесі внутрішньоклітинного дихання відбувається
аеробне окиснення органічних речовин (аеробне дихання). Енергія, яка при
цьому вивільняється, акумулюється у молекулі аденозинтрифосфату
(АТФ). АТФ є універсальною макроергічною сполукою.
Потім ця енергія використовується для перебігу процесів життєдіяльності
клітини – поділу, синтезу, поглинання та виділення речовин тощо. Мітохондрії
здатні активно рухатись. Вони концентруються навколо ядра, хлоропластів та
інших органел, де життєві процеси проходять дуже інтенсивно.
Мітохондрії мають власний спадковий апарат і власний білоксинтезуючий
апарат, завдяки чому можуть синтезувати білки. Вважають, що мітохондрії
можуть утворюватись двома шляхами: з ініціальних
часточок, відокремлених від ядра, та поділом мітохондрій. Пластиди характерні лише для рослин. Це морфологічно
відособлені білково – нуклеїново – ліпідні тільця,
здатні утворювати і зберігати пігменти, які зумовлюють забарвлення листків,
плодів, пагонів, квіток тощо. Залежно від наявності в них певних пігментів
розрізняють три типи пластид: безбарвні – лейкопласти, зелені – хлоропласти,
кольорові – хромопласти. Вони відрізняються не тільки забарвленням, а й
функцією. Досі не існує
єдиної думки щодо походження і утворення пластид. Так, є три гіпотези щодо їх
утворення: з вже існуючих; з ядра, мітохондрій та інших органел клітини;
шляхом новоутворення. Багато учених віддають перевагу гіпотезі генетичної
безперервності утворення пластид. Хлоропласти – пластиди зеленого кольору. Їх колір зумовлений
наявністю зеленого пігменту хлорофілу.
Саме з хлорофілом пов’язана основна функція хлоропластів – здійснення фотосинтезу – утворення органічних
речовин з вуглекислого газу і води за допомогою світлової енергії. Тому
хлоропласти є лише в клітинах надземних органів, на які потрапляє сонячне
світло, переважно в клітинах асиміляційної паренхіми листків – основних
органів, у яких відбувається фотосинтез. Значна кількість зелених пластид
міститься в поверхневих шарах клітин молодих пагонів, недозрілих плодів, корі
стебел, деревині, серцевинних променях та інших неглибоко розташованих
клітинах деревних і чагарникових рослин. Хлоропласти зустрічаються також у
сім’ядолях насіння (клен, гарбуз), ендоспермі насінин (хвойні), додаткових коренях (кукурудза). Кількість
хлоропластів у клітинах різна: у квасолі, огірків, рицини – 30–40, цукрового буряку, редьки – 50–60. У вищих рослин
хлоропласти здебільшого мають лінзоподібну форму. Діаметр їх – 4–6 мкм, товщина – 1–3 мкм. У
водоростей хлоропласти мають дуже різноманітну форму – зірчасту,
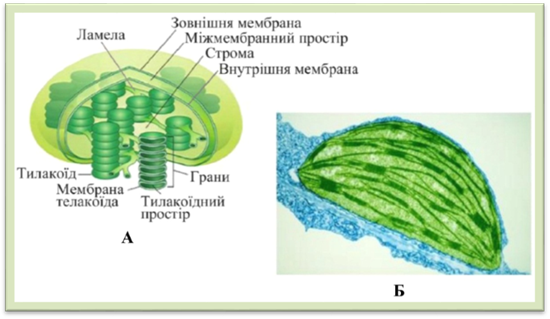
стрічкоподібну, сітчасту тощо. Оболонка
хлоропласта складається з двох мембран, між якими є міжмембранний
простір (рис. 1.1.7.8). Вона
містить великі й дрібні пори, крізь які сполучаються цитоплазма із
хлоропластом. Оболонка бере активну участь в обміні речовин. Крізь неї
проникають вода, амінокислоти, органічні й неорганічні фосфати, вона сприяє світлоіндукційному проникненню променів, газообміну,
дифузійному проникненню речовин, регулює швидкість метаболічних процесів.
Усередині хлоропластів міститься однорідна речовина – строма, пронизана системою паралельно розташованих
фотосинтетичних мембран двох типів. Одні з них у формі дисків, їх називають тилакоїдами.
Порожнина всередині тилакоїда називається тилакоїдним простором або люменом. Розміщуючись один над одним, тилакоїди
утворюють грани. Другі утворюють мембрани
різної конфігурації. Це ламели, або тилакоїди строми.
Грани зв’язані між собою тилакоїдами строми. У
хлоропластах різних рослин різна кількість гран: у клітинах мезофілу листка
шпинату – 40–60 гран, в інших рослин – до 200. Розмір гран 0,3–1,7 мкм. У кожній грані поєднується 3–60 тилакоїдів.
У тилакоїдах містяться основні фотосинтетичні
пігменти – хлорофіли, допоміжні пігменти –
каротиноїди та всі ферменти, необхідні для здійснення фотосинтезу. В стромі містяться
молекули ДНК, рибосоми, краплини ліпідів (їх називають пластоглобулами), крохмальні
зерна та інші включення.
Рис.
1.1.7.8. Хлоропласт: А – схема будови, Б – фотографія зроблена за
допомогою електронного мікроскопа Лейкопласти – безбарвні пластиди, в яких накопичуються запасні
поживні речовини. Містяться в клітинах насіння, бульб (рис. 1.1.7.9),
кореневищ, коренів. Вони прозорі, мають більшу щільність і світлозаломлення,
ніж цитоплазма. Звичайно, вони мають кулясту форму. Якщо ж в їх стромі
відкладається крохмаль або інші тверді сполуки, то вони набувають овальної,
паличкоподібної або нитчастої форми. Це найдрібніші з пластид, їх розміри не
перевищують 4 мкм. Внутрішня мембранна система
розвинена значно слабше, ніж у хлоропластів. У стромі є молекули ДНК,
рибосоми, пластоглобули.
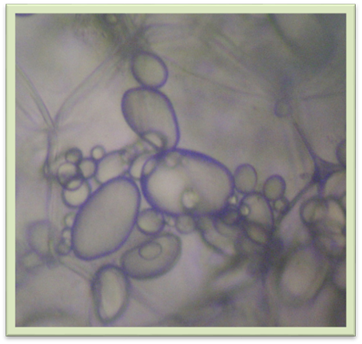
Рис. 1.1.7.9. Лейкопласти у клітині бульби картоплі Лейкопласти
виконують важливу роль у клітині. Синтезовані в листках речовини надходять до
лейкопластів і перетворюються за допомогою ферментів у різні форми запасних
поживних речовин. З цукрів, що надходять із фотосинтезуючих
органів, утворюються крохмальні зерна, різні за формою та розмірами, – вторинний
крохмаль. Запасний білок може відкладатись у вигляді кристалів або аморфних
гранул, олія – у вигляді пластоглобул. Лейкопласти, в
яких накопичується крохмаль, називаються амілопластами,
відкладаються про запас білки – протопластами,
жири – олеопластами,
нагромаджується про запас клітинний сік – гідропластами. Хромопласти – пластиди жовтого, оранжевого, червоного,
червонувато-коричневого та інших кольорів, які обумовлюють наявність різних (понад
60) пігментів – каротиноїдів. Дуже поширені каротин і ксантофіл, рідше
зустрічаються лікопін, лютеїн, родоксантин,
зеаксантин, криптоксантин. Хромопласти
зустрічаються (рис. 1.1.7.10)
у пелюстках квіток, плодах, насінні, є у вегетативних органах мохів, хвощів, паразитичних рослин. Зрідка бувають у
коренях.
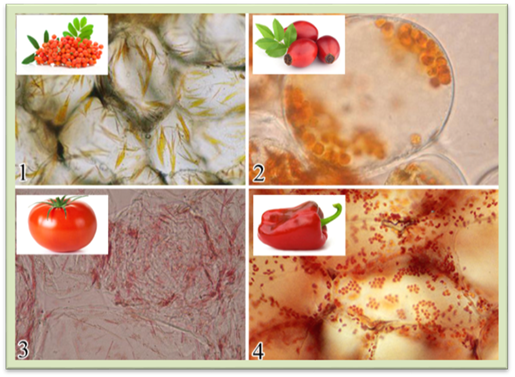
Рис.
1.1.7.10. Хромопласти у клітинах: 1 – горобини, 2 –
шипшини, 3 – томатів, 4 – червоного перцю Розміри
хромопластів різні; одні не перевищують 2–4 мкм,
інші досягають 10–12 мкм. Розміри пластид
змінюються залежно від інтенсивності відкладання в стромі білків, крохмалю,
краплин жиру. Внутрішньої
мембранної системи у них найчастіше немає. За формою
хромопласти дуже різноманітні: паличко–, голчасто–,
еліпсоподібні, лопатеві, трикутні тощо. Часто форма хромопластів залежить від
форми кристалів, що виникають внаслідок кристалізації каротину. Кристали
займають більшу частину строми і тим самим визначають форму пластид. Залежно
від форми накопичення каротиноїдів розрізняють хромопласти глобулярного, фібрилярного (трубчастого) і кристалічного типів. У хромопластів найпоширенішого глобулярного
типу пігменти розчинені в пластоглобулах. У
хромопластів фібрилярного типу каротиноїди не лише
наявні в пластоглобулах, а й утворюють групи
паралельних ниток або трубок, розташованих у стромі. У хлоропластів
кристалічного типу каротиноїди є в основному у вигляді кристалів різної
форми, які визначають форму самої пластиди (серпо-,
ромбо-, голкоподібну тощо). Хромопласти,
входячи до складу пелюсток квітів, є сигналізаторами для комах, що здійснюють
перехресне запилення. Наявність їх у плодах і насінні приваблює тварин, які
їх поїдають і розселяють. Хромопласти є джерелом провітаміну А. Пластиди здатні до взаємоперетворень. Розвиваються пластиди з пропластид – безбарвних тілець кулястої або амебоїдної форми розміром 0,4–0,9 мкм.
В їх будові розрізняють двомембранну оболонку,
зернистий матрикс, поодинокі елементарні мембрани,
невеликі крохмальні зерна, пластоглобули, пухирці. Пластиди одного типу здатні перетворюватися
на пластиди іншого. Так, на світлі в пропластидах
формується внутрішня система мембран, синтезується хлорофіл і вони
перетворюються на хлоропласти, а в глибині стебла і підземних органах – безбарвні лейкопласти. Те саме характерно і для лейкопластів, які здатні
перетворюватися на хлоропласти або хромопласти. Під час старіння листків,
стебел, дозрівання плодів у хлоропластах може руйнуватися хлорофіл,
спрощується будова внутрішньої мембранної системи і вони перетворюються на
хромопласти. Хромопласти є кінцевим етапом розвитку пластид: на пластиди
інших типів вони не перетворюються (рис. 1.1.7.11).
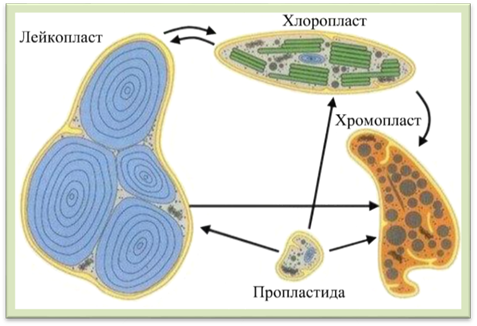
Рис.
1.1.7.11. Схема взаємоперетворення пластид 1.1.8. Фізіологічно активні
речовини цитоплазми До цієї групи
речовин належать: ферменти, вітаміни, фітогормони, антибіотики
та фітонциди. Ферменти – це клітинні каталізатори білкової природи, які прискорюють біохімічні
реакції. За складом ферменти поділяються на дві групи: однокомпонентні (прості),
які складаються з білків та двокомпонентні
(складні), які складаються з
білкової та небілкової частин, яка може бути представлена похідними
вітамінів, або катіонами чи аніонами. Каталітичну
функцію виконує ділянка ферменту – активний
центр. Його структура відповідає будові речовин, які вступають у реакцію.
Тому ферменти характеризуються високою специфічністю, тобто певний фермент каталізує лише один тип реакцій. Так, інвертаза
(сахараза) розщеплює сахарозу на глюкозу і фруктозу. В той же час цей фермент
не діє на інші сполуки. Активність
ферментів залежить від температури, рН середовища,
тиску, концентрації субстрату й самого ферменту. Відомо близько
2000 ферментів, які за характером їхньої дії поділяють на шість класів: оксидоредуктази
(каталізують реакції
окиснення, тобто перенесення електронів
з однієї молекули на іншу), трансферази (каталізують перенесення функціональних груп і
молекулярних залишків від однієї молекули до іншої; гідролази (каталізують
розщеплення за участю води складних органічних сполук
на прості), ліази
(каталізують реакції негідролітичного відщеплення
від субстрату і приєднання певних груп атомів з утворенням подвійних зв’язків, а також зворотні реакції), ізомерази
(каталізують перетворення органічних сполук на їхні ізомери внаслідок внутрішньомолекулярних
переміщень атомів), лігази
(каталізують з'єднання
двох молекул з утворенням нового хімічного зв'язку). Вітаміни – це фізіологічно-активні речовини, які необхідні для здійснення
нормальних фізіологічних процесів у тварин і людини. Вітаміни відкрито М.
Луніним у 1880 р., а назву їм дав польський учений К. Функ
у 1912 р. Зелені рослини – основний постачальник продуктів харчування,
багатих на вітаміни. Тепер відомо понад 40 вітамінів, які об’єднуються у дві
групи: жиророзчинні (А, Д, Е, К) та
водорозчинні (В, С, Н, РР). Вітамін А. У рослинах містяться провітаміни A (попередники вітаміну) – каротиноїди.
Найбільш важливим серед них є каротин. Багато його у моркві, шпинаті, зеленій цибулі, плодах шипшини, горобини,
малини, томатів, червоного солодкого перцю. З
каротину в організмі людини і тварин синтезується вітамін А. При нестачі
вітаміну А в організмі затримується ріст, знижується стійкість до
захворювань, виникає захворювання очей (куряча сліпота), уражуються тканини
епітелію. Вітаміни групи D відіграють важливу роль у метаболізмі кальцію і
фосфору. Містяться у дріжджах, пліснявих грибах, олії льону, соняшнику, кукурудзи. При нестачі цих
вітамінів діти та молодняк ряду тварин хворіють рахітом. Найбільш активними
формами цього вітаміну є ергокальциферолу (вітамін D2), що
надходить в організм тільки з їжею та холекальциферолу
(вітамін D3), який синтезується у
шкірі під дією ультрафіолетових променів. Вітамін Е впливає на статеву функцію тварин та людей, його нестача викликає
деградацію статевих залоз, м’язову дистрофію, атрофію зародка. Він
зустрічається в томатах, цитрусових, гороху, салаті, петрушці, пшениці, олії льону і бавовнику, а також у тваринних
продуктах. Вітамін К бере участь в регуляції процесів зсіданні крові, сприяє
зміцненню капілярів та припиненню кровотеч. Міститься у листках каштану, кропиви, люцерни, білоголовій капусті, щавлі, шпинаті, петрушці, томатах. Вітамін В1 (тіамін) у
великій кількості накопичується пшеничних і рисових висівках, дріжджах, картоплі, гречці. Нестача його в
організмі є причиною порушень діяльності нервової системи, множинного
ураження нервів у людини і тварин. Вітамін В2 (рибофлавін)
міститься у дріжджах, гречаній та вівсяній
крупі, зеленому горошку, шпинаті, моркві, картоплі тощо. Нестача цього
вітаміну спричинює зниження гостроти зору, трофічної функції, центральної
нервової системи, погіршує стан шкіри та слизових оболонок і кровотворну
функцію кісткового мозку. Вітамін В6 (піридоксин)
міститься у дріжджах, пшеничних і рисових висівках, пшеничних зародках, волоських горіхах і фундуку, в шпинаті, картоплі, моркві, цвітній і білоголовій капусті, помідорах, полуниці, черешні, апельсинах і лимонах. При його нестачі
виникають дерматити, анемія, порушується білковий обмін і синтез жирів Вітамін В12 (ціанокобаламін) бере участь у кровотворенні, регулює
вуглеводний і жировий обмін в організмі. При його недостатності розвивається
недокрів'я – анемія. Цей вітамін відсутній в рослинних продуктах, але його
багато в продуктах тваринного походження – яловичій печінці, нирках, серці. Вітамін С (аскорбінова кислота)
зустрічається в цибулі, зеленому горошку, капусті, картоплі, кропі, перці, хвої, мандаринах, шипшині, смородині, недозрілому волоському горіху, лимонах тощо. Нестача вітаміну
С викликає цингу, знижує стійкість організму до зовнішніх негативних впливів
та інфекційних захворювань. Вітамін Н (біотин) у малих кількостях
міститься у всіх продуктах, але більше всього цього вітаміну міститься в дріжджах, бобових (соя), цвітній капусті, арахісі. У меншій мірі він
міститься в томатах, шпинаті, в грибах. Здорова мікрофлора
кишечника синтезує біотин в достатній для організму кількості. Його нестача
спричиняє ураження шкіри, нігтів, випадання волосся, слабкість м’язів,
сонливість, анемію тощо. Вітамін РР (нікотинамід) у великих
кількостях міститься в дріжджах, висівках, пшениці, пшеничних зародках, кукурудзі, картоплі. Нестача нікотинаміду
в організмі є причиною захворювання на пелагру, що супроводжується ураженням
шкіри, психічними розладами. Фітогормони – це фізіологічно активні речовини, що виробляються спеціальними
тканинами рослин і діють як регулятори і координатори онтогенезу. Серед них є стимулятори (ауксини, гібереліни, цитокініни) і інгібітори
(абсцизова кислота, етилен). Перші здатні
прискорювати або підсилювати певні процеси в організмі, другі – сповільнюють
перебіг хімічних реакцій, а також біохімічних і фізіологічних процесів. Ауксини продукуються верхівками
пагонів і коренів. Вони стимулюють розтягнення клітин і ріст органів,
сприяють утворенню коренів на живцях. Гібереліни
викликають прискорення росту і цвітіння рослин, партенокарпію
(розвиток без насіннєвих плодів). Цитокініни активують поділ клітин. Абсцизова кислота затримує
поділ і розтягнення клітин, гальмує ріст, прискорює листопад, затримує
проростання насіння. Етилен
прискорює дозрівання і старіння всіх частин рослин. Антибіотики – це речовини, що виробляються клітинами мікроорганізмів і вбивають або
пригнічують ріст інших мікроорганізмів. Антибіотики є продуктами
життєдіяльності деяких грибів і широко застосовуються як лікувальні засоби в
боротьбі з хвороботворними мікроорганізмами. Зараз вивчено близько 500
антибіотиків. У 1940 р. О. Флемінг і його колеги вперше виділили антибіотик пеніцилін з пліснявих грибів Penicillium notatum.
Він є засобом для боротьби з мікроорганізмами, що викликають пневмонію,
газову гангрену, його застосовують при лікуванні гнійно-септичних та
запальних процесів. Стрептоміцин виробляється
ґрунтовими актиноміцетами Actinomyces streptomycini. Застосовується для лікування
туберкульозу, бруцельозу, туляремії, коклюшу тощо. Левоміцетин виділений з культури актиноміцету Actinomyces
venezuelae. Застосовується для боротьби з висипним
і черевним тифом, гострими кишковими інфекціями. Тетрацикліни є продуктом
життєдіяльності актиноміцетів: окситетрациклін – Streptomyces rimosus, хлортетрациклін – Streptomyces
aureofaciens. Застосовуються як стимулятори
росту в кормових добавках у тваринництві. Антибіотики застосовуються і в рослинництві.
Так, антиміцин, який пригнічує ріст деяких
грибів, застосовується для боротьби з грибковими захворюваннями цибулі, гороху, рису, винограду. Фітонциди – органічні речовини, що утворюються клітинами вищих рослин і
пригнічують або згубно діють на мікроорганізми. Відкрив цю групу речовин Б. Токін у 1928 р. Вони захищаються рослини від руйнівної
дії мікроорганізмів. Так, жито виділяє бензоксазолінон, який захищає
його від ураження грибом фузаріумом, горох виділяє ізатин, який згубно діє на ряд грибів,
що спричиняють грибкові захворювання. Відомими фітонцидами є аліцин,
виділений з цибулі, сативін – з часнику, рафінін – з редьки і редиски, кварцетин – з дуба, крепін – з скереди покрівельної, іманін – зі звіробою. Тривалість їхньої
дії – від 50 до 500 годин і більше. Фітонциди використовують для лікування
респіраторних захворювань, гнійних ран, для профілактики грипу, захворювань
травного каналу. 1.1.9. Ядро, його структура, роль Ядро
найважливіший і обов’язковий компонент еукаріотичної клітини. Воно є місцем збереження й відтворення спадкової інформації,
яка визначає ознаки цієї клітини і всього організму в цілому, а також є
центром керування синтезу білків. Лише деякі клітини втрачають ядро під
час свого розвитку (ситоподібні трубки рослин). Більшість клітин має одне
ядро, в окремих грибів і водоростей –
два і більше. У прокаріотичних організмів (бактерії, синьо-зелені водорості) немає сформованого
ядра, а речовини, які входять до його складу, містяться у цитоплазмі. Ядро функціонує
лише у цитоплазматичному середовищі. Кожному типу клітин властиве постійне
співвідношення між об’ємом ядра й цитоплазми (ядерно-цитоплазматичне співвідношення). Порушення цих
співвідношень або спричинює поділ клітини, або призводить до їх загибелі.
Так, у молодих клітин співвідношення між об’ємами ядра і всього протопласта
дорівнює від 1:4 до 1:5, в у зовсім сформованих і у старих – від 1:20 до
1:200. Розмір, форма, положення ядра
постійні, але можуть змінюватися залежно від віку, фізіологічного стану і
впливу екологічних факторів. Ембріональні клітини мають великі ядра,
розміщені в центрі, а зрілі – менші по відношенню до об'єму клітини і зміщені
в пристінний шар. Ядро може зміщуватися рухом цитоплазми. Форма ядер
різноманітна, але, як правило, вона відповідає формі клітини: у паренхімних
клітинах ядро найчастіше кулеподібне, в прозенхімних
– лінзо- або веретеноподібне. Розміри ядер
різні: у грибів їх діаметр 0,5–1,0 мкм, квіткових
рослин – 5–25, саговників – до 500 мкм, середній
розмір 10–25 мкм. У структурі ядра розрізняють: ядерну оболонку, каріоплазму, ядерце, хроматин (хромосоми) (рис. 1.1.9.1).
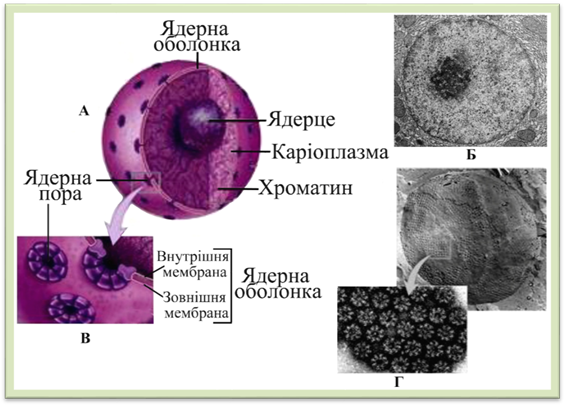
Рис.
1.1.9.1. Будова ядра: А – схема будови, Б – фотографія ядра зроблена за
допомогою електронного мікроскопа; В – схема будови ядерної оболонки і ядерних пор, Г
– мікрофотографія ядра зроблена за допомогою сканувального
електронного мікроскопа (помітні ядерні пори) Ядерна оболонка відмежує вміст ядра від цитоплазми. Вона просторово і функціонально з’єднана із мембранами ендоплазматичної сітки (рис. 1.1.9.2). Складається з двох
мембран – зовнішньої та внутрішньої, між якими
є перинуклеарний
простір, заповнений матриксом,
який за своєю структурою та хімічним складом нагадує гіалоплазму цитоплазми. Товщина мембран 10 нм, а товщина перинуклеарного
простору варіює. Загальна товщина оболонки 40–80 нм. У більшості клітин під
час поділу ядерні мембрани зникають і знову утворюються після завершення
поділу. За структурою і
хімічним складом ядерна оболонка подібна до ендоплазматичної сітки. Внутрішня
мембрана оболонки агранулярна, до зовнішньої
мембрани прикріплені рибосоми. Зовнішня мембрана утворює вирости, які переходять у ретикулум цитоплазми.
Рис.
1.1.9.2. Функціональний зв’язок ядерної оболонки з гранулярною
ендоплазматичною сіткою Ядерна оболонка
має особливі утворення – ядерні пори (рис. 1.1.9.1), навколо яких зовнішня і внутрішня мембрани сполучені між собою. Через
пори ядро сполучається з цитоплазмою. Діаметр пори 80-90 нм. Це складні структури. По межі
пори, що утворена внаслідок злиття двох мембран, розташовані гранули, від
яких відходять фібрили. Частина фібрил збігається у центрі, формуючи
діафрагму порового комплексу. Завдяки ядерній оболонці між
цитоплазмою і ядром виникає взаємозв'язок. Ядерна оболонка контролює обмін речовин між ядром і
цитоплазмою, крізь ядерні пори макромолекули рухаються з ядра у гіалоплазму і
у зворотному напрямку. Ядерний сік, або каріоплазма, або нуклеоплазма
являє собою колоїдний розчин, в якому розташовані одне або кілька ядерець,
хроматин і різноманітні гранули. Каріоплазма неоднорідна за консистенцією і
нагадує надосадову рідину і в цьому відношенні
подібна до гіалоплазми. Залежно від фізіологічного
стану вона набуває форми
золя або геля. В'язкість її порівняно низька. В каріоплазмі містяться прості білки (гістони, протаміни) і високомолекулярні
(нуклеопротеїди, які включають ДНК, РНК, аскорбінову
кислоту, мінеральні солі кальцію), є численні ферменти. В ній не лише відбувається зв'язок між структурами ядра, а
й трансформуються речовини,
які рухаються по ній. Ядерце. В ядрі одне або кілька ядерець, рідше їх багато. Ядерце не постійне,
воно зникає на початку і з'являється вкінці поділу клітини. За формою
кулясте. Розміри його невеликі і часто коливаються залежно від розмірів ядра,
тривалості інтерфази, умов живлення. Об'єми ядра і ядерця співвідносяться в меристемних клітинах як 16:1, а в клітинах постійних
тканин – 40:1. Ядерце складається
в основному з білка та РНК . За фізичними
властивостями ядерце подібне до ядра, прозоре, напіврідкої консистенції,
більш щільної, ніж каріоплазма. Щільну субмікроскопічну структуру ядерець
створюють паралельно розміщені білкові фібрилярні
нитки, які називають нуклеонемами,
і гранули. Знаходяться вони в безструктурному аморфному матриксі,
подібному до гіалоплазми. Ядерце контактує з
вторинною перетяжкою хромосоми, яку називають організатором ядерця. На ній відбувається матричний синтез рибосомної РНК (рРНК). Потім рРНК сполучається з білком, внаслідок чого утворюються
гранули рибонуклеопротеїдів – субодиниці рибосом,
які потрапляють в нуклеоплазму і крізь пори ядерної
оболонки проникають у цитоплазму, де з’єднуються у рибосоми. Отже, функцією ядерець є синтез рибосомної РНК (рРНК), а також утворення субодиниць
рибосом. Хроматин – ниткоподібні структури ядра, утворені здебільшого з
білків та нуклеїнових кислот. Ділянки хроматину неоднорідні. Ті з них, що
постійно перебувають в ущільненому стані, називають гетерохроматином. Вони добре
забарвлюються різними барвниками і в період між поділами клітини помітні у
світловий мікроскоп. Незабарвлені, менш ущільнені ділянки мають назву еухроматин. На початку поділу
клітини нитки хроматину ущільнюються і з них
формуються компактні тільця – хромосоми.
Під кінець клітинного поділу хромосоми розкручуються до ниток
хроматину. Хромосоми – основні структури ядра, які забезпечують
збереження і передачу спадкової інформації дочірнім клітинам у процесі
клітинного поділу. Хромосоми можуть перебувати у двох станах. У робочому –
це деконденсовані
тонкі (10 нм) нитчасті структури, що беруть активну участь у процесі обміну
речовин, їх видно лише під електронним мікроскопом. Під час поділу ядра
хромосоми максимально конденсуються,
стають короткими і товстими, їх видно під світловим мікроскопом. Виконують
функцію розподілу й перенесення генетичної інформації. За хімічною природою хромосома являє собою нуклеопротеїд, що складається з ДНК та білка. Крім того,
до складу хромосом входять РНК та ферменти. Ядерні білки утворюють особливі
структури – нуклеосоми,
кожна з яких складається з восьми білкових глобул. Особливі білки зв’язують нуклеосоми між собою. Молекули ДНК у хромосомах накручені
навколо нуклеусом. Така організація забезпечує компактне розміщення молекул
ДНК у хромосомах, оскільки довжина цих молекул у розгорнутому стані значно
перевищує довжину хромосом (рис.
1.1.9.3). Хромосоми досягають 0,2-50 мкм в
довжину і 0,2–2 мкм в товщину.
Рис.
1.1.9.3. Організація хромосоми Хромосома
складається (рис. 1.1.9.4) з
двох поздовжніх частин – хроматид, кожна
з яких містить по молекулі ДНК з подібним набором спадкової інформації. Кожна
хромосома складається з двох хроматид, а хроматида – з двох спіралізованих хромонем. В свою
чергу хромонему утворює пучок елементарних ниток,
що є основою субмікроскопічної будови хромосоми. Елементарна нитка
утворюється з ДНК, основного білка і невеликої кількості кислих білків. На
певних ділянках хромонем знаходяться хромомери, які занурені в матрикс
хромосом. Хромосома має первинну перетяжку (неконденсована ділянка), яка поділяє
хромосому на плечі. У зоні первинної перетяжки є ділянка, яка з’єднує хроматиди, – центромера. На ній
формуються білкові структури – кінетохори. Під час поділу клітини до кінетохора приєднуються нитки веретена поділу і
забезпечують розподіл хроматид або цілих хромосом
між дочірніми клітинами. Кінці плечей хромосоми називають теломерами,
це спеціалізовані ділянки, які перешкоджають з'єднанню хромосом між собою або
з їхніми фрагментами.
Рис. 1.1.9.4. Будова хромосоми: А – схема будови, Б –
фотографія хромосоми зроблена за допомогою електронного мікроскопа Хромосоми відрізняються розмірами, положенням первинної перетяжки,
довжиною плечей по обидва боки центромери.
Хромосоми, в яких перинна перетяжка знаходиться посередині хромосоми і плечі
рівної або майже однакової довжини, називають метацентричні (рівноплечі).
Якщо перетяжка зміщена від середини і одне плече довше за інше – субметацентричні (малонерівноплечі) хромосоми. Акроцентричні
(різконеріноплечі)
хромосоми – це хромосоми, в яких перетяжка розміщена близько до одного
із кінців і одне плече довше, а друге – дуже коротке і часто малопомітне. Хромосоми, у яких
залишилось тільки одне плече з центромерою на кінці
в результаті відриву другого плеча, називаються телоцентричні (паличкоподібна). В
нормальному наборі хромосом такі хромосоми не зустрічаються. Деякі хромосоми
мають ще й вторинну перетяжку, що
функціонує як ядерцевий організатор. Вона відокремлює від
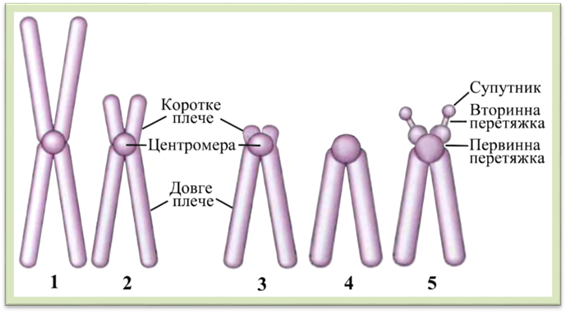
хромосоми невеликий фрагмент – супутник (рис. 1.1.9.5).
Рис.
1.1.9.5. Типи хромосом: 1 – метацентрична, 2 – субметацентрична, 3 – акроцентричні,
4 – телоцентрична, 5 – супутникова Хромосоми є постійними і обов'язковими для ядра. Для кожного виду рослин
кількість в клітинах стала. Їх сукупність у клітині утворює хромосомний набір
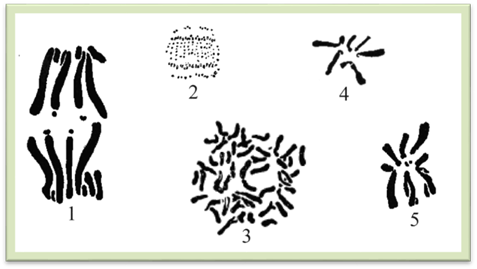
– каріотип (рис. 1.1.9.6). Хромосомний набір
ядра може бути гаплоїдним,
диплоїдним або поліплоїдним. У гаплоїдному наборі (1n) усі хромосоми відрізняються одна від іншої за будовою. Гаплоїдний набір хромосом властивий статевим клітинам,
спорам, клітинам мохоподібних. Натомість у диплоїдному наборі (2n)
кожна хромосома має парну, подібну за розмірами та особливостями будови. Всі
соматичні клітини рослини, які розвиваються із зиготи, є диплоїдними.
Диплоїдний набір хромосом цибулі становить 16, жита – 14, кукурудзи – 20, лілії – 24. Хромосоми однієї пари називають гомологічними. Відповідно хромосоми,
які не належать до певної пари, негомологічні
одна до одної. Якщо ж кількість гомологічних хромосом перевищує дві, то такі
хромосомні набори належать до поліплоїдних: триплоїдний (3n),
тетраплоїдний (4n) тощо.
Рис.
1.1.9.6. Каріотипи рослин: 1 – алое, 2 – липи, 3 –
картоплі, 4 – буряку, 5 – скерди 1.1.10. Поділ ядра і
клітини як основа процесів розмноження і розвитку рослин Ріст і розвиток рослин тісно пов'язаний
з новоутворенням клітин, завдяки чому утворюються різні тканини, вегетативні
та генеративні органи рослин. Появі нових клітин передує поділ ядра.
Найпоширеніші три способи поділу ядра і клітини: амітоз; мітоз, або
каріокінез; мейоз, або редукційний поділ. Завдяки поділу ядра і клітини
відбувається ріст і розмноження клітин. Багаторазовий поділ клітин сприяє
наростанню органічної маси за рахунок галуження коренів і пагонів, утворення
великої кількості однойменних органів. Таким чином збільшується всисна
поверхня коренів і листків, наростають бульби картоплі, коренеплоди буряку, кореневища пирію, яблука, зернівки пшениці тощо. Однак досі ще
повністю не з'ясовано причини багаторазового поділу клітин окремих рослин або
органів. Поки що неможливо пояснити, чому, наприклад, внаслідок
багаторазового поділу клітин виникає невеличка рослинка ряски в кілька
міліметрів завбільшки, або ж гігантський евкаліпт висотою
понад 150 м. Безумовно, ріст і поділ клітин регулюють ДНК, але поки що
процес саморегуляції активності ДНК ще не цілком зрозумілий. Питання
регулювання ДНК цікавить багатьох вчених і його розв’язання дозволить
вирішувати теоретичні і практичні проблеми сільськогосподарського
виробництва. 1.1.11. Поняття про мітотичний цикл. Інтерфаза Мітотичний
цикл – це період існування клітини, який починається з
моменту її утворення шляхом поділу материнської клітини, і закінчується
власним поділом. Мітотичний цикл включає два періоди:
інтерфазу і мітоз (рис. 1.1.11.1).
Рис. 1.1.11.1. Періоди мітотичного циклу Інтерфаза – це період між двома послідовними поділами клітини або від завершення
останнього поділу до загибелі клітини. В інтерфазі клітина росте, синтезує органічні
сполуки, запасає енергію у вигляді макроергічних сполук та готується до поділу. Тривалість інтерфази
становить 90% часу всього клітинного циклу. В інтерфазі розрізняють три
послідовні періоди: пресинтетичний(G1),
синтетичний (S), постсинтетичний (G2). Пресинтетичний
період (G1) настає зразу після закінчення поділу. Тут
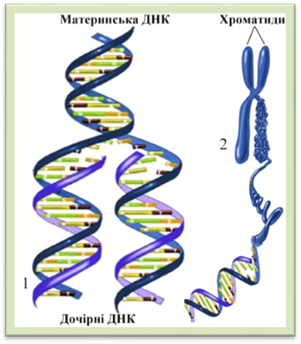
відбувається біосинтез білка та всіх видів РНК, відтворюються мембранні структури та органели. Клітина інтенсивно росте і може виконувати свою функцію. У синтетичному періоді (S) відбувається синтез ДНК (реплікація) (рис.1.1.11.2). На кожному з ланцюгів деспіралізованої молекули ДНК добудовується
комплементарний ланцюг. На кінець періоду кількість молекул ДНК подвоюється.
Кожна хромосома
складається з двох хроматид, які містить по молекулі ДНК з подібним набором
спадкової інформації. Продовжується синтез РНК і білків. Постсинтетичний період (G2)
відзначається інтенсивним синтезом білків, РНК і накопиченням енергії.
Рис. 1.1.11.2. Реплікація ДНК (1); двохроматидна
хромосома (2) 1.1.12. Мітоз. Цитокінез Кожному типу
клітин властиве певне стале співвідношення між об’ємами ядра та цитоплазми
(ядерно-цитоплазматичне співвідношення). Внаслідок росту клітини порушується
ядерно-цитоплазматичне співвідношення, що призводить до поділу клітини. Мітоз (від грец. mitos – нитка), або
каріокінез, або непрямий поділ – це основний спосіб поділу еукаріотичних соматичних
клітин. Процес мітозу супроводжується спіралізацією хромосом та
утворенням особливого апарату, який забезпечує рівномірний розподіл
спадкового матеріалу материнської клітини між двома дочірніми. Мітоз
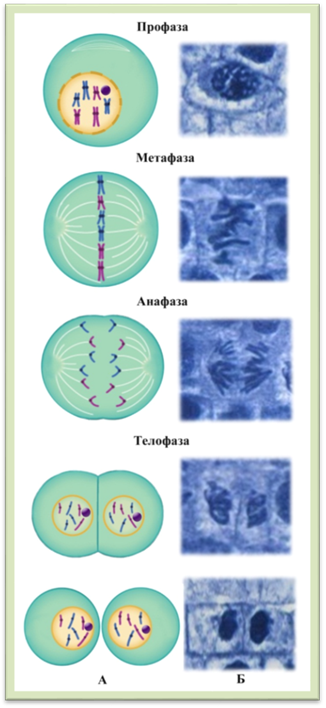
включає чотири послідовні фази (рис. 1.1.12.1): профазу, метафазу, анафазу,
телофазу.
Профаза
починається з ущільнення ниток хроматину: хроматиди вкорочуються і потовщуються (спаралізуються).
Поступово руйнуються ядерця, ядерна оболонка розпадається, внаслідок чого хромосоми потрапляють
до цитоплазми. Починає формуватись веретено поділу. Метафаза
характеризується завершенням спіралізації хромосом і формуванням веретена
поділу. Хромосоми розташовуються в площині екватора, утворюючи метафазну пластинку. Кожна хромосома поділяється на дві хроматиди, які з’єднані в ділянці центромери,
якою хромосома прикріплюється до однієї з ниток веретена поділу. Анафаза – це
найкоротша із фаз, під час якої нитки веретена скорочуються, внаслідок чого
зв'язок між хроматидами руйнується і вони в якості
самостійних хромосом рухаються до протилежних полюсів клітини, руйнується
веретено поділу. Після завершення руху, на полюсах утворюються два рівноцінні
набори хромосом. Телофаза триває від
завершення розходження хроматид до утворення
дочірніх клітин. На початку телофази хромосоми деспіралізуються.
На кожному полюсі навколо хромосом утворюється ядерна оболонка, формуються
ядерця і клітина стає двоядерною. Потім цитоплазма
поділяється на дві рівні частини (цитокінез), утворюючи дві однакові дочірні клітини. Цитокінез починається в кінці телофази з
потовщення ахроматинових ниток, що призводить до
утворення серединної пластинки, яка від центру до периферії розростається і
поділяє протопласт материнської клітини на дві дочірні. Кожна з них має
сформовану клітинну оболонку, цитоплазму, ядро, пластиди та інші органели. Біологічне
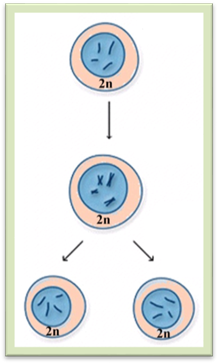
значення мітозу (рис. 1.1.12.2) полягає в тому, що мітоз забезпечує точну
передачу спадкової інформації від материнської клітини дочірнім. Кожна
дочірня клітина одержує по одній хроматиді від кожної
материнської хромосоми, внаслідок
чого в усіх дочірніх клітинах зберігається стала кількість хромосом протягом
послідовних клітинних циклів.
Рис. 1.1.12.2. Схема, що ілюструє значення мітозу
Мітоз у рослинній клітині Амітоз, або прямий
поділ, мало поширений в природі, але добре відомий в деяких нижчих і вищих рослин, в клітинах конуса наростання стебла і кореня, в паренхімі черешків
листків, нуцелусі, ендоспермі, клітинах стінок зав'язі,а також в дуже старих і хворих
клітинах з яких дуже часто розвиваються ракові пухлини у рослин (рис. 1.1.13.1). Суть амітозу полягає в тому, що ядро
без попередньої підготовки подовжується, перетягується і ділиться на два
дочірні. При цьому ядерна оболонка не зникає і не виникають нові хромосоми і хромосомний
апарат. Кожне з ядер одержує різну кількість ДНК, а
значить вони різні не тільки за вмістом нуклеїнової
кислоти, а й за формою, розмірами, життєвістю.
Рис. 1.1.13.1. Рак у рослин: 1 – на бульбах картоплі; 2 – звичайний рак троянд;3 – чорний рак яблуні У результаті поділу ядер окремих рослин виникають не дві, а 4–8 або й більше різновеликих або рівних ядер.
Такий спосіб утворення ядер називають
фрагментарним. Амітоз відбувається природним шляхом
або може бути викликаний штучно: під впливом випромінювання, а також
факторів, які порушують хід мітотичного поділу. Мейоз – спосіб
поділу клітин, у результаті якого кількість хромосом зменшується вдвічі й
клітини переходять з диплоїдного стану в гаплоїдний.
Мейоз властивий рослинам, які розмножуються безстатевим і статевим способами.
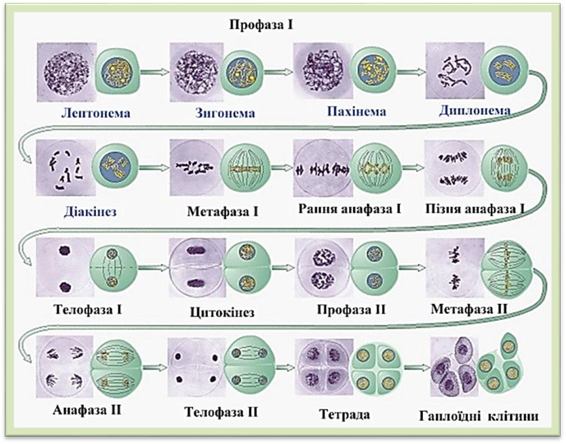
Мейоз відбувається перед утворенням пилку та зародкового мішка у квіткових, перед утворенням спор у мохів, хвощів, голонасінних. У процесі мейозу виділяються два послідовні поділи: редукційний і екваційний. Кожний з цих поділів, як і мітоз, складається з чотирьох послідовних фаз (рис.1.1.14.1).
До початку мейозу в інтерфазі відбувається
подвоєння
кількості
ДНК і кожна
хромосома
стає
двохроматидною.
І
редукційний поділ мейозу.
Профаза
І більш
тривала
і складніша,
ніж
інші.
В ній виділяються п'ять стадій: лептонему, зигонему, пахінему, диплонему, діакінез. Для лептонеми властива поява двох довгих
і тонких хроматинових ниток з хромомерами – хромосоми. Поодинокі хромосоми, які називаються моновалентами, досягають високої деспіралізації. Зигонема відзначається попарним зближенням і кон'югацією двох гомологічних хромосом, моновалентів,
тобто утворенням бівалентів, які відображають процес кон'югації хромосом. Пахінема характеризується вкороченням і потовщенням хромосом внаслідок їх подальшої спіралізації. Гомологічні хромосоми редуплікують (подвоюються). На місці бівалентів виникають чотири хроматиди. Згодом гомологічні хромосоми в результаті
кросинговеру обмінюються гомологічними ділянками (рис.
1.1.14.2).
Рис. 1.1.14.2. Схема кросинговеру: 1 – гомологічні хромосоми утворюють
біваленти; 2 – обмін ділянками
між гомологічними хромосомами (кросинговер); 3 –
гомологічні хромосоми після кросинговеру. Диплонема в зв'язку з подальшою спіралізацією
призводить до ще більшого потовщення хромосом. Гомологічні хромосоми в
окремих місцях розходяться і залишаються з'єднаними в місцях перехрещення. Так
виникають хіазми. Діакінез - це
подальша спіралізація, вкорочення і потовщення хромосом. Ядерце зменшується в
розмірах. У метафазі
І розчиняються ядерна оболонка і ядерце, виникають ахроматинові нитки. Новоутворені
біваленти розміщуються в центрі, створюючи своєрідну пухку екваторіальну
площину. В анафазі
І гомологічні хромосоми розходяться до протилежних полюсів, відбувається
редукція хромосом. Телофаза І
відзначається утворенням двох ядер, але без поділу
хромосом. Виниклі з диплоїдної клітини ядра мають гаплоїдний
(1n) набір
хромосом. Інтерфаза між першим та
другим мейотичними поділами вкорочена або відсутня
у більшості рослин. Молекули ДНК у цей період
не подвоюються. ІІ екваційний поділ мейозу. Екваційний поділ
починається фактично з метафази, оскільки в телофазі попереднього поділу
виниклі ядра та їх хромосоми далі не діляться, а перебувають в стані спокою. Екваційний поділ відбувається подібно до того, як при
мітозі. В метафазі
ІІ моноваленти,
що складаються з двох хроматид, розміщуються в
екваторіальній площині і прикріплюються кінетохорами
до нових ахроматинових ниток. В анафазі
ІІ хроматиди
розходяться до протилежних полюсів і набувають розмірів материнських
хромосом. Для телофази ІІ характерні утворення дочірніх ядер
з оболонкою і ядерцем, деспіралізація хромосом, а
також формування поперечної перетинки. Отже, внаслідок першого мейотичного поділу виникають клітини або лише ядра з гаплоїдним набором хромосом. У результаті другого мейотичного поділу число хромосом залишається таким, як і
після першого, але двохроматидні хромосоми
розділяються на однохроматидні і кількість ДНК,
унаслідок розходження хроматид до дочірніх клітин,
зменшується вдвічі. Біологічне значення мейозу. В результаті мейозу з однієї диплоїдної материнської
клітини утворюються чотири (тетрада) гаплоїдні дочірні клітини (рис. 1.1.14.3). Значення мейозу не лише в забезпеченні
кількості хромосом у організмі з покоління в покоління, тобто збереження
сталості виду. Завдяки випадковому розподілу гомологічних хромосом і обміну
їх окремих ділянок в мейозі гаплоїдні статеві
клітини, що утворилися внаслідок цього поділу, містять найрізноманітніший
збіг хромосом від статевих клітин попереднього покоління. Це забезпечує
різноманітність хромосомних наборів і відповідно ознак у наступних поколінь
і, таким чином, дає матеріал для еволюції організмів.
Рис. 1.1.14.3. Схема, що ілюструє значення мейозу
Мейоз у рослинній клітині 1.1.15. Порушення процесу мейозу. Поліплоїдія У деяких випадках утворенню статевих
клітин не передує зменшення кількості їхніх хромосом, і вони залишаються диплоїдними.
В результаті цього всі клітини рослини, що сформувалися після запліднення, будуть мати три (якщо редукція відбулася тільки у
одного партнера), чотири, а іноді і більше наборів хромосом. Такі клітини і
рослини, що мають ці клітини, називають поліплоїдними.
Виникнення поліплоїдності може бути і
наслідком порушень у мейозі, які зумовлені штучно (опроміненням, обробкою
хімічними речовинами та ін.) Поліплоїди мають дещо
інший вигляд, ніж диплоїди (рис.1.1.15.1). Вони більші за розміром, більш облистнені, мають великі квітки, більш врожайні
(рис.1.1.15.2).
Рис. 1.1.15.1. Зовнішній вигляд троянд: 1 – диплоїдної; 2 – поліплоїдної
Рис. 1.1.15.2. Квіти поліплоїдних рослин: 1 – лілійника; 2 – шафрана; 3 – іриса Багато високопродуктивних сортів
культурних рослин (помідорів, кукурудзи, пшениці, буряків та ін.) є поліпоїдами (рис.1.1.15.3).
Рис. 1.1.15.3. Поліплоїдні рослини: 1 – триплоїдний кавун;
2 – тетраплоїдна малина; 3 – тетраплоїдні
кормові буряки В результаті порушень з тих або інших
причин мітозу поліплоїдними можуть стати окремі
клітини або тканини тієї самої рослини. Запитання і завдання для самоконтролю 1. Що таке мітотичний цикл? З яких періодів він
складається? 2. Що таке інтерфаза? Яке її значення для існування
клітини? 3. Які існують типи поділу клітин? 4. Що таке мітоз? У чому полягає біологічне значення
мітозу? 5. Охарактеризуйте кожну фазу мітозу. 6. Що таке мейоз? Яке значення мейозу? З яких поділів
він складається? 7. Що таке кон’югація і кросинговер гомологічних
хромосом? Яке значення кросинговеру? 8. Що
спільного та відмінного між процесами мітозу та мейозу? |
|||