|
|
ЕПІЗООТОЛОГІЯ З МІКРОБІОЛОГІЄЮ I частина Електронний посібник |
||||||||
|
|
10. Основи загальної вірусології |
||||||||
|
10.1. Морфологія і фізіологія вірусів. 10.1.3. Хімічний склад вірусів 10.4. Методи
лабораторної діагностики вірусних інфекцій 10.4.2. Методи ідентифікації вірусів 10.5. Віруси бактерій
(бактеріофаги) 10.1. Морфологія і фізіологія вірусів Відомо тисячі
видів різноманітних вірусів людини, тварин, комах, рослин, бактерій. Вони
відіграють надзвичайно важливу роль у природі: виступають як фактор, що
об’єднує складні живі системи органічного світу, служать переносниками
генетичної інформації. Саме за допомогою вірусів зроблені фундаментальні
відкриття з розшифрування структури нуклеїнових кислот, механізмів
реплікації ДНК та синтезу білка. Фундаментальне вивчення вірусів та
бактеріофагів увінчалося грандіозними успіхами генної інженерії. Отже, вони
дають ключ до розуміння функціонування нуклеїнових кислот і сутності життя. Віруси – неклітинні форми живих істот, які характеризуються
малими розмірами, відсутністю власних білоксинтезуючих та енергієгенеруючих
систем, а також облігатним внутрішньоклітинним
паразитизмом.
Окрема
вірусна частинка одержала назву віріон. Він складається з однієї молекули
нуклеїнової кислоти (ДНК або РНК) та білкового футляра, що її оточує, –
капсида. Разом вони формують нуклеокапсид. Капсиди утворені з білкових
субодиниць (поліпептидів), які називаються капсомерами. Їх кількість
стабільна для кожного виду вірусів і використовується як таксономічна ознака.
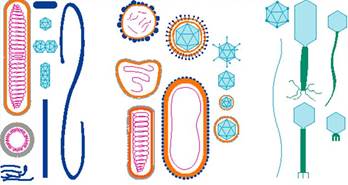
Віруси з таким типом будови називають простими. До них
належать найдрібніші з патогенних вірусів: поліовіруси, аденовіруси,
паповавіруси. Проте
більшість вірусів має ще одну оболонку – суперкапсидну, яка містить ліпіди.
Вона пронизана вірусспецифічними білками-глікопротеїдами, які на поверхні
оболонки утворюють особливі стуктури, що називаються шипами. Такі віруси
називають складними або оболонковими. Структурною одиницею суперкапсидної
оболонки є пепломер. До них належать віруси сказу, герпесу, грипу, енцефалітів
та ін. Віріони
характеризуються поняттям симетрії. Тип симетрії залежить від способу укладки
нуклеїнової кислоти і, відповідно, розташування капсомерів навколо неї.
Виділяють ізометричний (або кубічний), спіральний та змішаний типи симетрії.
Кубічний тип характеризується тим, що капсомери утворюють багатогранник
(найчастіше ікосаедр – 20-гранник). У віріоні зі
спіральною симетрією молекула нуклеїнової кислоти закручена разом із
капсомерами в тугу спіраль. Такий тип симетрії мають віруси мозаїчної хвороби
тютюну, грипу, кору, епідемічного паротиту та ін. Комбінований
тип симетрії спостерігається у деяких бактеріофагів. При цьому головка
бактеріофага має кубічний тип симетрії, а нуклеопротеїд, розміщений у хвості,
укладається спірально.
В основі сучасної
класифікації вірусів лежать ознаки, що характеризують тип нуклеїнової
кислоти, їх морфологію, особливості репродукції, антигенні властивості, чутливість
до фізичних і хімічних агентів, особливо ефіру, імунологічні властивості,
природні механізми передачі, патологія, формування включень, симптоматологія
захворювань. За наявністю
нуклеїнової кислоти їх поділяють на ДНК-геномні та РНК-геномні
віруси. Із 71 відомої родини вірусів 20 родин містять віруси, патогенні для
людини і тварини. Визначення
розмірів вірусів здійснюють фільтруванням, з допомогою ультрацентрифугування
та спостереження в електронному мікроскопі. Порівняльні розміри: (1) Staphylococcus має діаметр 1000 nm, (2) Бактеріофаги
–10–100 nm, (3) Діаметр молекули сироваткового альбуміну 5 nm, глобуліну – 7
nm. 10.1.3. Хімічний склад вірусів Віруси
містять лише один тип нуклеїнової кислоти (ДНК або РНК), яка становить від 1
до 40 % маси віріона. Вірусні геноми містять інформацію, достатню для синтезу
лише декількох білків. Їх маса сягає 10–15 мкг, що в 1 млн разів менше, ніж у
клітини, а довжина – до 0,093 мкм. Число нуклеотидних пар коливається від
3150 (вірус гепатиту В) до 230000 (вірус натуральної віспи). Віруси
характеризуються надзвичайним розмаїттям форм геному. Він може бути
представлений як односпіральними, так і двоспіральними молекулами, бути
лінійним, циркулярним або фрагментованим. Білки вірусів
(70–90 % маси віріона) поділяються на структурні та
неструктурні. Структурними називають такі білки, які
входять до складу зрілих позаклітинних віріонів. Вони виконують низку
важливих функцій: захищають нуклеїнову кислоту від зовнішнього пошкодження,
взаємодіють з мембранами чутливих клітин і забезпечують проникнення вірусу в
клітину. Неструктурні білки не входять до складу зрілих віріонів, однак
утворюються під час їх репродукції. Вони забезпечують регуляцію експресії
вірусного геному, є попередниками вірусних білків, здатні пригнічувати
клітинний біосинтез. Залежно від розташування у віріоні, білки поділяються на
капсидні, суперкапсидні, матриксні, білки серцевини та асоційовані з
нуклеїновою кислотою. Різні групи
вірусів мають неоднакову стійкість до дії факторів зовнішнього середовища.
Наприклад, віруси грипу, парагрипу, епідемічного паротиту інактивуються на
поверхнях за декілька годин, проте аденовіруси зберігають інфекційні
властивості декілька днів. Чутливість вірусів до дії рентгенівського та
ультрафіолетового опромінення залежить від величини геному вірусів: чим менший геном, тим резистентніший вірус
до опромінення. Віруси, які мають ліпопротеїдну оболонку, чутливі до ефіру,
хлороформу та дезоксихолату натрію, інших жиророзчинників і детергентів. Важливою
особливістю вірусів є їх чутливість до концентрації водневих іонів. Частина з
них стійка до кислих значень рН (2,2–3,0). До них
належать віруси, які викликають кишкові інфекції, проникаючи в організм
аліментарним шляхом. Віруси, які потрапляють в організм через верхні дихальні
шляхи (риновіруси, віруси грипу та ін.), чутливі до кислих значень рН.
Завдяки
сучасним методам дослідження вченим вдалося встановити складний характер
взаємодії вірусу з клітиною хазяїна. Залежно від властивостей вірусу і
сприйнятливої клітини, а також умов зовнішнього середовища, розрізняють такі
основні типи взаємодії вірусу з
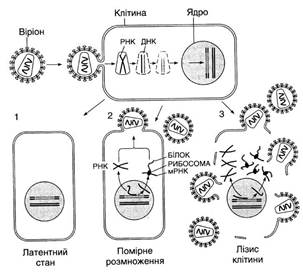
клітиною: а) розмноження вірусу призводить до руйнування і загибелі клітини
(такий тип взаємодії дістав назву продуктивної інфекції, а віруси –
вірулентних); б) взаємодія, під час якої клітини залишаються живими, а
дозрілі вібріони не утворюються, називається абортивною інфекцією; в) геном
вірусу інтегрується (об'єднується) з генетичним апаратом клітини і під час
поділу клітини може тривалий час передаватися в дочірні клітини. Однак через
деякий час, за певних умов, у клітині може початися розмноження вірусу, яке
призводить клітину до загибелі. Такий тип взаємодії дістав назву вірогенії. Репродукція
вірусів – унікальна система відтворення
чужерідної інформації в клітинах еукаріотів і забезпечує абсолютне підкорення
клітинних структур потребам вірусів. У репродукції
вірусів виділяють низку стадій. До ранніх –
належить адсорбція вірусів на поверхні клітини, проникнення (пенетрація) їх
всередину кілтини та їх роздягання (депротеїнізація). Пізні стадії
(стратегія вірусного генома) включають синтез вірусних нуклеїнових кислот,
синтез білка, збирання віріонів та вихід вірусних часток із клітини. Прості віруси
залишають клітину, як правило, шляхом «вибуху», розриваючи її мембрану. Складні
віруси – брунькуванням. При цьому клітина тривалий час може залишатись
життєздатною, поки повністю не виснажиться, продукуючи вірусних нащадків. 10.4. Методи
лабораторної діагностики вірусних інфекцій Лабораторна
діагностика вірусних інфекцій може здійснюватись за двома основними
напрямками. Перший – виявлення та ідентифікація
вірусів або їх компонентів в організмі хворого (вірусологічна діагностика).
Другий напрямок – виявлення специфічних противірусних антитіл у сироватці крові (серологічна діагностика). Оскільки застосування
цього методу практично підтверджує перенесення захворювання, його ще
називають ретроспективною діагностикою. Практично для всіх вірусних інфекцій
реакції, які використовуються з цією метою, вважаються позитивними, якщо в
досліді виявлено чотирикратний і більше приріст титру антитіл.
Для виділення
вірусів, необхідних для подальшого їх дослідження з метою ідентифікації, дуже
важливо провести правильний забір, швидке транспортування матеріалу до
лабораторії, раціональний вибір тест-системи (курячі ембріони, культури
клітин, лабораторні тварини). Однак у багатьох випадках досліджуваний
матеріал може містити бактеріальну флору, яка під час подальшого зараження
тест-систем спотворюватиме результати досліджень, включаючи їх загибель. Тому
в лабораторіях використовують методи знищення бактерій у доставлених зразках.
Надійним методом є обробка матеріалу антибіотиками, а простих вірусів –
ефіром. Допустимим вважається використання спеціальних антибактеріальних
фільтрів або центрифугування матеріалу під час невисоких швидкостей. У
цьому випадку бактерії опускаються з осадом на дно. Для
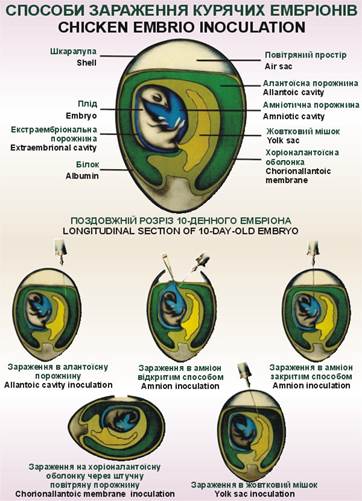
розмноження вірусу необхідна наявність живих чутливих клітин. Курячі ембріони
6–12-денного віку є зручною моделлю для виділення та подальшої ідентифікації
вірусів. Після зараження їх інкубують у термостаті за температури 36–38 °С упродовж
декількох днів. Для виділення
вірусів у курячих ембріонах їх можна заражати на хоріоналантоїсну оболонку, в
алантоїсну порожнину, порожнину амніону, жовтковий мішок тощо. Численні
лабораторні тварини широко використовуються у вірусології для виділення та
ідентифікації вірусів, отримання специфічних противірусних сироваток,
вивчення різноманітних аспектів патогенезу вірусних захворювань, розробки
способів боротьби із захворюваннями та їх профілактики. Найчастіше
використовують білих мишей різного віку (навіть одно- і дводенного), білих
щурів, гвінейських свинок, кролів, ховрахів, бавовникових щурів, мавп та
інших. Існують
різноманітні способи зараження тварин залежно від тропізму вірусів, клінічної
картини захворювання тощо. Досліджуваний матеріал можна вводити через рот, у
дихальні шляхи (інгаляційно, через ніс), нашкірно, внутрішньошкірно, підшкірно,
внутрішньом’язово, внутрішньовенно, внутрішньоочеревинно, внутрішньосерцево,
на скарифіковану рогівку, у передню камеру ока, у мозок. Репродукція
вірусів у культурах клітин (культури ембріонів людини, нирок мавп,
фібробластів ембріона курки тощо ) використовується для лабораторної
діагностики вірусних інфекцій. Цьому сприяла розробка методів культивування
клітин в умовах in vitro і створення штучних живильних середовищ для них,
відкриття антибіотиків, які використовуються для пригнічення розмноження
сторонньої мікрофлори тощо. Живильні
середовища, які використовуються для підтримання культур клітин або їх росту,
бувають природними або синтетичними (штучними). Природні середовища –
сироватка крові великої рогатої худоби, рідини із серозних порожнин, продукти
гідролізу молока, різноманітні гідролізати (5 %
гемогідролізат, 0,5 % гідролізат лактальбуміну) або екстракти тканин.
Залежно від
функціонального використання середовища можуть бути ростові (з великим
вмістом сироватки крові) – їх використовують для вирощування клітинних
культур до внесення вірусних проб, і підтримуючі (з меншим вмістом сироватки
чи її відсутністю) – для утримання інфікованих вірусом клітинних культур. Внаслідок
розмноження вірусів у культурі клітин у них виникають дегенеративні зміни, які
позначаються як цитопатична дія вірусів (ЦПД). Виявляти віруси в культурі
тканин можна за феноменом бляшкоутворення. За морфологічними змінами в
культурі клітин можна виділити декілька типів прояву цитопатичної дії. Її
особливості дозволяють провести попередню діагностику захворювання. Досить часто як
прояв цитопатичної дії віруси утворюють внутрішньоклітинні включення. Вони
можуть локалізуватись як внутрішньоядерно, так і в цитоплазмі клітин. Їх
утворення, форма, величина, наявність у них вірусних нуклеїнових кислот є
важливим моментом під час проведення вірусологічної діагностики захворювань.
Такі включення утворюють віруси сказу (тільця Бабеша-Негрі), натуральної віспи (тільця Гварнієрі), простого
герпесу (тільця Ліпшютца), аденовіруси, віруси грипу та ін. Включення
виявляють під час світлової мікроскопії, фарбуючи препарати за допомогою
методу Романовського-Гімзи. Часто для виявлення включень використовують метод
імунофлуоресценції. В ультрафіолетових променях люмінесцентного мікроскопа
включення світяться характерним кольором. Можна довести наявність включень у
матеріалі (біоптати) за допомогою електронної мікроскопії, яка дозволяє
дослідити тонку структуру цих утворень, виявити окремі віріони. Часто для
виявлення внутрішньоядерних або внутрішньоцитоплазматичних включень
використовують відбитки тканин або органів, зскрібки клітин або гістологічні
зрізи із тканин загиблих. 10.4.2. Методи ідентифікації вірусів Важливим
етапом лабораторної діагностики будь-якої вірусної інфекції є виявлення та
типування вірусів у досліджуваному матеріалі, яке передбачає використання
специфічних противірусних сироваток. Реакція
гальмування гемаглютинації (РГГА) базується на здатності блокувати
гемаглютинуючі властивості вірусів за допомогою специфічних антитіл. У
результаті цього спостерігається затримка аглютинації еритроцитів.
Реакція
гальмування гемадсорбції базується на тому, що еритроцити набувають здатності
адсорбуватись на поверхні культур клітин, які зазнали модифікації внаслідок
появи в оболонці глікопротеїнів вірусів.
Антитіла імунної сироватки під час взаємодії з поверхневими антигенами
вірусів, представленими в оболонці, зв’язують їх, внаслідок чого гальмується
адсорбція еритроцитів на клітинах. Реакція
нейтралізації ґрунтується на взаємодії специфічних антитіл імунної сироватки з
вірусом, яке призводить до нейтралізації останнього. Кольорова
проба передбачає, що під час взаємодії вірусів з культурою клітин останні
гинуть, рН середовища залишається лужним, і колір індикатора не змінюється.
За нейтралізації вірусів антитілами специфічної сироватки або за відсутності
відповідних вірусів створюються умови для розвитку культур клітин. Вони
залишаються живими, активно здійснюють обмін речовин, внаслідок чого
утворюються продукти клітинного метаболізму, які зсувають рН середовища в
кислу сторону. Це призводить до зміни кольору індикатора фенолроту з
червоного (лужне рН) на солом’яно-жовтий або оранжевий (кисле значення рН). Реакцію
зв’язування комплементу досить широко використовують у вірусологічній
практиці для ідентифікації хвороботворних агентів. Вона належить до непрямих
двосистемних гетерологічних реакцій і ґрунтується на принципі взаємодії
комплементу із специфічним комплексом антиген-антитіло. Внаслідок цього не
відбувається гемолізу сенсибілізованих еритроцитів. У реакціях
імунодифузії використовують принцип дифундування антигенів і антитіл
назустріч один одному в напіврідкому середовищі, наприклад, агаровому гелі. У
зоні оптимальних співвідношень антигенів та антитіл утворюються лінії
преципітації. Їх число залежить від кількості антигенів, які відрізняються
своїми епітопами, тому що кожна лінія відповідає своїй системі антиген –
антитіло. Метод
флуоресціюючих антитіл (МФА) набув широкого застосування у вірусології внаслідок
високої специфічності та зручності в користуванні. Існують численні
модифікації цього методу. Зокрема, можна виявляти та ідентифікувати віруси як
безпосередньо в досліджуваному матеріалі, так і після попереднього зараження
ними, наприклад, культур клітин. Принцип методу полягає у взаємодії антигенів
з антитілами, які заздалегідь мічені флуорохромами (родаміном, який дає люмінесценцію червоного кольору, або
флуоресцеїнізотіоцианатом натрію, який зумовлює зелене світіння комплексу в
ультрафіолетових променях). Метод непрямої імунофлуоресценції можна ефективно
використовувати для ідентифікації більшості вірусів. Імуноферментні
методи широко використовуються у лабораторній практиці; особливо під час
імуногістологічних досліджень, а також для виявлення циркулюючих антигенів,
антитіл та імунних комплексів, які мають суттєве значення в діагностиці
інфекційних хвороб. 10.5. Віруси бактерій
(бактеріофаги) Природним
місцем існування фагів є бактеріальна клітина, тому фаги поширені всюди
(наприклад, у стічних водах). Фагам притаманні біологічні особливості, які
властиві і іншим вірусам. Найбільш
морфологічно поширений тип фагів характеризується наявністю
голівки-ікосаедра, відростка (хвоста) зі спіральною симетрією (часто має
порожнистий стрижень і скорочувальний чохол), шипів і відростків (ниток). Взаємодія
фагів з клітиною (бактерією) суворо специфічна, тобто бактеріофаги здатні
інфікувати тільки певні види бактерій. За спектром
дії фаги поділяють на: - полівалентні (лізують близькоспоріднені
бактерії, наприклад сальмонели); - моновалентні (лізують бактерії одного
виду); - типоспецифічні (лізуют тільки з певного
збудника). Одержують бактеріофаги або з лізогенних культур мікроорганізмів, або з
довкілля, заражаючи матеріалом відповідні бактерії. Для цього досліджуваний
матеріал центрифугують, для осадження твердих частинок, а надосадову рідину в
об’ємі 1 мл вносять у 30 мл м’ясо-пептонного бульйону, який попередньо
засіяний 1 мл 6–18-годинної культури бактерій. Через 18–24 год інкубації за
температури 37 °С посів фільтрують через бактеріальний фільтр, розводять у
співвідношення 10:1, 10:2, 10:3, 10:4 разів, додають 0,05–0,1 мл культури
мікроорганізмів та інкубують упродовж доби за оптимальної температури. Як
контроль використовують культуру бактерій без бактеріофагу. Якщо в матеріалі
присутній бактеріофаг, середовище залишається прозорим внаслідок лізису
бактерій фагами, в той час як у контрольній пробірці відбувається ріст
бактерій (середовище стає мутним). Віруси 3. Яку функцію виконують структурні білки вірусу? 4. В чому особливість
репродукції вірусів? 5. Назвіть два основних напрямки
діагностики вірусних інфекцій? 6. Що собою являє реакція гемаглютинації? |
|||||||||